Accepted Version
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
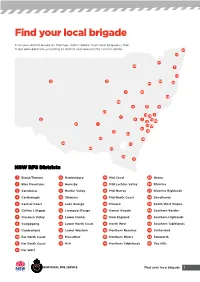
Find Your Local Brigade
Find your local brigade Find your district based on the map and list below. Each local brigade is then listed alphabetically according to district and relevant fire control centre. 10 33 34 29 7 27 12 31 30 44 20 4 18 24 35 8 15 19 25 13 5 3 45 21 6 2 14 9 32 23 1 22 43 41 39 16 42 36 38 26 17 40 37 28 11 NSW RFS Districts 1 Bland/Temora 13 Hawkesbury 24 Mid Coast 35 Orana 2 Blue Mountains 14 Hornsby 25 Mid Lachlan Valley 36 Riverina 3 Canobolas 15 Hunter Valley 26 Mid Murray 37 Riverina Highlands 4 Castlereagh 16 Illawarra 27 Mid North Coast 38 Shoalhaven 5 Central Coast 17 Lake George 28 Monaro 39 South West Slopes 6 Chifley Lithgow 18 Liverpool Range 29 Namoi Gwydir 40 Southern Border 7 Clarence Valley 19 Lower Hunter 30 New England 41 Southern Highlands 8 Cudgegong 20 Lower North Coast 31 North West 42 Southern Tablelands 9 Cumberland 21 Lower Western 32 Northern Beaches 43 Sutherland 10 Far North Coast 22 Macarthur 33 Northern Rivers 44 Tamworth 11 Far South Coast 23 MIA 34 Northern Tablelands 45 The Hills 12 Far West Find your local brigade 1 Find your local brigade 1 Bland/Temora Springdale Kings Plains – Blayney Tara – Bectric Lyndhurst – Blayney Bland FCC Thanowring Mandurama Alleena Millthorpe Back Creek – Bland 2 Blue Mountains Neville Barmedman Blue Mountains FCC Newbridge Bland Creek Bell Panuara – Burnt Yards Blow Clear – Wamboyne Blackheath / Mt Victoria Tallwood Calleen – Girral Blaxland Cabonne FCD Clear Ridge Blue Mtns Group Support Baldry Gubbata Bullaburra Bocobra Kikiora-Anona Faulconbridge Boomey Kildary Glenbrook -

Bland Temora Zone Bush Fire Management Committee Bush Fire Risk Management Plan 2020
Bland Temora Zone Bush Fire Management Committee Bush Fire Risk Management Plan 2020 Table of Contents Glossary 7 Chapter 1. Introduction 9 1.1 Background 9 1.2 Aim and Objectives 9 1.3 Description of the Bland Temora BFMC Area 10 1.3.1 Location and land tenure 10 1.3.2 Climate and bush fire season 10 1.3.3 Population and demographic information 10 1.3.4 History of bush fire frequency and ignition cause 11 Chapter 2. Identifying and Assessing the Bush Fire Risk 12 2.1 Process 12 2.2 Communication and Consultation 12 2.3 Identifying the Bush Fire Risk 13 2.3.1 Assets 13 2.3.2 Assessing the Bush Fire Risk - Consequence 14 2.3.3 Assessing the Bush Fire Risk - Likelihood 15 2.3.4 Identifying the level of risk 15 2.3.5 Evaluating the Bush Fire Risk 15 2.3.6 Prioritising Treatments 15 2.3.7 Risk Acceptability 16 Chapter 3. Treating the Risk 17 3.1 Bush Fire Management Zones 17 3.2 BFMC Wide Treatments 19 3.3 Asset Specific Treatments 20 3.4 Fire Thresholds 21 3.5 Annual Works Programs 22 3.6 Implementation 23 Chapter 4. Performance Monitoring and Reviewing 24 4.1 Review 24 4.2 Monitoring 24 4.3 Reporting 24 4.4 Performance Measurements 24 Appendix 1 Community Participation Strategy 25 Appendix 2 Asset Register 25 Appendix 3 Treatment Register 25 Appendix 4 Maps 25 Page 2 of 34 List of Tables Table 1.1 Land tenure 10 Table 3.1 Bush Fire Management Zones: Purpose, objectives and characteristics 18 Table 3.2 Asset specific treatments used in the (insert name here) BFMC area 21 Table 3.3 Fire Thresholds for Vegetation Categories 22 List of Figures Figure 2.1 Overview of the risk assessment process ................................................................................... -

25Th APRIL 2016
25th APRIL 2016 Commemorative booklet proudly presented by Michael McCormack MP Federal Member for Riverina Assistant Minister for Defence WAR-TIME TOTS: A World War I postcard of an Australian patriotic scene showing two young girls dressed as nurses carrying a stretcher. WAR TOUGH ON CHILDREN OUR poignant cover of this year’s commemorative the south to the Bland and the South West Slopes in the ANZAC Day Riverina booklet features a caring nurse north, the Riverina has always contributed mightily in lovingly tending to a brave little trooper, wounded in battle. the nation’s darkest days. PITCHING IN: When it was war-time a woman’s work Role-playing the heroic deeds being done by the grown- War cost the Riverina heavily yet has never dented the was never done for if they weren’t nursing at the front, local ups at the front was one way children of The Great resolve of the region’s people to serve and to play their ladies were keeping the home fires burning and busy fund War era – indeed, littlies during any time of military part – at home or abroad. raising to help those fighting the good fight for God, King and conflict – were able to cope with the horrors of what was And now – as we mark a century since that terrible year CONTENTS Country. happening half a world away. of 1916 in which so many of our volunteer Diggers fell INSIDE: Playing dress-ups with siblings and friends was harmless at the killing fields of Bullecourt, Fromelles, Messines, and popular fun. -

Junee Roadside Vegetation Survey and Management Guidelines
CONTENTS FOREWORD........................................................................................................ 2 ACKNOWLEDGEMENTS .............................................................................................. 3 INTRODUCTION 1.1 The importance of roadside vegetation.................................................. 5 1.2 The Junee Roadside Vegetation Survey................................................ 6 1.3 Responsibility for road reserve management......................................... 7 1.4 Junee Shire – an overview..................................................................... 8 1.5 Survey Methodology .............................................................................. 8 SURVEY RESULTS 2.1 Roadside vegetation in Junee Shire .................................................... 12 2.2 Species Recorded................................................................................ 12 2.3 Native Plants........................................................................................ 14 2.4 Introduced Plants ................................................................................. 18 MANAGEMENT OF ROADSIDE VEGETATION 3.1 Management according to conservation status.................................... 21 3.2 Recommendations to Council .............................................................. 22 REFERENCES ...............................................................................................................25 APPENDICES A Junee Shire roadside vegetation data .......................................................26 -

Southern NSW Research Results 2015
Southern NSW research results 2015 RESEARCH & DEVELOPMENT – INDEPENDENT RESEARCH FOR INDUSTRY www.dpi.nsw.gov.au Southern NSW research results 2015 RESEARCH & DEVELOPMENT – INDEPENDENT RESEARCH FOR INDUSTRY an initiative of Southern Cropping Systems Editors: Deb Slinger, Director Southern Cropping, NSW DPI, Wagga Wagga; Elizabeth Madden Disclaimer and Cynthia Podmore, NSW DPI, Wagga Wagga; The information contained in this publication is based on and Carey Martin, NSW DPI, Orange. knowledge and understanding at the time of writing (May 2016). However, because of advances in knowledge, users are reminded Reviewer: Don McCaffery, Technical Specialist of the need to ensure that the information upon which they rely Pulses & Oilseeds, NSW DPI, Orange. is up to date and to check the currency of the information with Cover images: Main image–lupin variety trial, the appropriate officer of NSW Department of Industry, Skills and Regional Development or the user’s independent adviser. Mark Richards, NSW DPI, Wagga Wagga; inset canola and wheat–NSW DPI Image Library; inset The product trade names in this publication are supplied on field pea–Di Holding, AnDi Communications. the understanding that no preference between equivalent products is intended and that the inclusion of a product ©State of NSW through NSW Department of Industry, name does not imply endorsement by the department Skills and Regional Development 2016 over any equivalent product from another manufacturer. ISBN 978-1-74256-930-7 Recognising that some of the information in this document jn 13880 is provided by third parties, the State of New South Wales, the author and the publisher take no responsibility for the Published by NSW Department of Primary Industries, a part of accuracy, currency, reliability or correctness of any information NSW Department of Industry, Skills and Regional Development included in the document provided by third parties. -

Michael Mccormack MP
Commemorative booklet proudly presented by Michael McCormack MP Federal Member for Riverina CONTENTS GALLIPOLI’S LASTING LEGACY GALLIPOLI is not just a place but initiative, respect and also now very much a condition in the teamwork. human spirit so profound it empowers Over the years Australians to be their best selves ... Australia’s brave and patriotic with a sense of the servicemen and importance of mateship burning women have always deeply within. put the interests of Although by military standards the maintaining and AWM WORTH VISITING 1915 ANZAC campaign was a disaster at times restoring - an epic one at that - the symbolism freedom - and the inherent risks THE Australian War of Gallipoli and what those hardy and associated with going on active duty - Memorial in Canberra holds heroic Diggers achieved truly united above their own personal safety. one of the world’s largest Australia and Australians like nothing collections of material related else could possibly have at the time. Long lines of crosses - some marked, to World War I. others not - in military cemeteries and Looking back now, we should not only be row upon row of names on the Roll of The redevelopment of The THE LANDING: Troops and horses go ashore at the Dardanelles. proud but also thankful, eternally thankful, Honour at the Australian War Memorial Great War galleries in time for the deeds which established the ethos and on monuments across the Riverina for the ANZAC Centenary which is held so dear by all who wear a are grim reminders that our nation has created an even greater INSIDE: military uniform of our country today. -

Junee NSW VCA Plant Communities
Office of Environment & Heritage Junee Native Vegetation Map NSW VCA Plant Communities ADS-40 Edition 2 NSW VCA ID NSW VCA Name Total Area - Landform pattern / main soil types Characteristic species in each stratum. Note that floristics are relevant to NSW VCA community over 1:100,000 (8428) this map its entire distribution, and may not accurately reflect community make-up within this mapsheet (ha) Grassy Woodlands Floodplain Transition Woodlands 70 White Cypress Pine woodland on sandy loams in central NSW 37 Peneplain, Plain / Red earth, Red-brown Callitris glaucophylla / Acacia deanei subsp. deanei - Dodonaea viscosa sens lat. - Maireana wheatbelt earth enchylaenoides - Geijera parviflora / Einadia nutans subsp. nutans - Austrostipa scabra subsp. scabra - Austrodanthonia eriantha - Sida corrugata 76 Inland Grey Box tall grassy woodland on alluvial loam and clay 3,302 Alluvial plain, Flood plain / Grey clay, Red Eucalyptus microcarpa / Dodonaea viscosa subsp. cuneata - Acacia buxifolia subsp. buxifolia / soils in the NSW South Western Slopes and Riverina Bioregions clay, Red-brown earth Austrodanthonia caespitosa - Austrostipa scabra subsp. falcata - Chloris truncata - Sida corrugata 80 Inland Grey Box - White Cypress Pine tall woodland on loam soil 529 Alluvial plain, Stagnant alluvial plain / Grey- Eucalyptus microcarpa - Callitris glaucophylla - Allocasuarina luehmannii / Maireana microphylla - Acacia 546000 547000 548000 549000 550000 551000 552000 553000 554000 555000 556000 557000 558000 559000 560000 561000 562000 563000 564000 565000 566000 567000 568000 569000 570000 571000 572000 573000 574000 575000 576000 577000 578000 579000 580000 58!1000 582000 583000 584000 585000 586000 587000 588000 589000 590000 591000 on alluvial plains of NSW Southwestern Slopes and Riverina brown podzolic soil, Red-brown earth deanei subsp. -

RESULTS of the BARRY FERGUSON CLASSIC 2015 Special Notes
RESULTS of the BARRY FERGUSON CLASSIC 2015 There were no radar penalties and I congratulate all crews on their behavior on the road. As all penalties for navigational issues were the same value, the results have been shown as the number of penalties without multiplying by 25. The Results for the Masters and Apprentices are on following sheets of this spreadshhet document. The Results for the event are shown on the sheet marked 'Results' The detailled results for each Division are seperated for Masters and Apprentices on subsequent sheets. These sheets identify the places where penalties were accrued. The reference to points referred to in the details are identified on the sheet marked Reference Boards' Special Notes. The reults have been calculated on the course as set except for the matters listed below. Sect 5 Divn B We had not stated a finish time for the days assembly and consequently closure of the Passage Control at Hanwood was incorrect. No penalties were attributed to the later cars in Division B for the points B30 and 31. Sect 7 Divn C Old and New Alignments of Burley Griffin Way. Kamarah to Ardlethan Due to the visibility of the false Board from the Correct Board at this location and the fact that there was no advice that VRCs will be on the left hand side of the road, the penalty for recording the false Board has been deleted when the correct board has also been recorded. Sect 7. Divn C Travel in Yarrangerry SF. The advice given of the distance that the Director had travelled was incorrect and while only 'Advice' and not an instruction, misled competitors . -

Inside DISABILITY 8 DIOCESE 26 TIMES 31
TOGETHERIn the Diocese of wagga.catholic.org.au FREE DECEMBER 2018 DAY OF PEOPLE SCHOOLS CHRISTMAS WITH A IN OUR MASS inside DISABILITY 8 DIOCESE 26 TIMES 31 O come let us adore him 2 FROM THE APOSTOLIC ADMINISTRATOR TOGETHER IN THE DIOCESE OF WAGGA WAGGA is published monthly, eleven Christmas 2018 - times per year, and is available free at parish churches, schools, presbyteries and religious houses throughout the Wagga Wagga we beg to differ! Diocese. Publisher The Diocese of Wagga Wagga Apostolic Administrator of the Wagga Wagga Diocese Archbishop Christopher Prowse - McAlroy House 205 Tarcutta Street Jesus. homeless ness. For most, will not allow human selfishness Wagga Wagga NSW 2650 His coming was prefigured by Christmas is a time of gatherings and aggression to have the last the Angel of light’s encounter for Church and family re say. With Jesus – never! There is Editor/Designer Karene Eggleton with the Shepherds. This unions. Sadly, for too many, the always mercy and forgiveness. heavenly light “stood over them thought of family gatherings is It is certainly true to say at Printer and the glory of the Lord shone a heartache and not a reason for Christmas – “Isn’t it great to be The Riverina Media Group Peter Street Wagga Wagga 2650 round them”. (Lk.2/9) joy. The darkness of loneliness loved!” But it is even greater to Here the cosmos or universe is arises in the hearts of some say “Isn’t it great to love!” Submissions com municating a language for marriages and families. Jesus is always found in TOGETHER would not be us all. -

Chlorophyll in Australian Canola
AUSTRALIAN OILSEEDS FEDERATION INCORPORATED FINAL REPORT "Chlorophyll in canola" AOF 15-1 1 May 2000 to 30 June 2002 Dr R.J. Mailer, Agricultural Research Institute, Wagga Wagga (02 69381818) ISBN 0 7347 1421 1 AOF chlorophyll final report 1 Fig 1. (Cover). Samples of canola oil from canola seed samples containing increasing levels of chlorophyll of 6, 23, 35, 53 and 81 mg/kg respectively. AOF chlorophyll final report 2 AUSTRALIAN OILSEEDS FEDERATION INCORPORATED R&D PROJECT 2000/2002 Project Title: Chlorophyll in canola Reference number given by AOF to Preliminary Proposal: AOF 15-1 Organisation: NSW Agriculture Administrative Contact: Title First name Surname Mr Graham Denney Address: NSW Agriculture, Locked Bag 21, ORANGE NSW 2800 Phone: (02) 6391 3219 Fax: (02) 6391 3327 Email: [email protected] Principal Investigator: Address: Title First name Surname Wagga Wagga Agricultural Institute, PMB, Wagga Dr Rodney Mailer Wagga, NSW 2650 Phone: 02 69381818 Fax: 02 69381809 Email: [email protected] Starting date # : 1 / 5 /2000 Finishing date # : 30 / 06 /2002 AOF chlorophyll final report 3 Project Outline and Budget Process Criterion Cost Total 1 Genotype by Analyse canola samples from site x 100 x $31 $3100 environmental cultivar x year. Approx 100 relationship samples. 2 BHC’s study Obtain samples from bulk handlers 100 x $31 $3100 and canola trials/crops with the worst levels of chlorophyll/green seed. Approx 100 samples. 3 Green seed Determine the relationship between 50 x $10 $1500 screening test number of green seeds and spectrophotometric reading. Say 50 samples tested for green seed by dip test and compared with spectrophotometric test. -
Carmelite Monastery of Our Lady Queen of Peace Wagga Wagga
TOGETHERIn the Diocese of wagga.catholic.org.au FREE NOVEMBER 2019 Mass of Thanksgiving Carmelite Monastery of Our Lady Queen of Peace 1966-2019 Wagga Wagga 2 FROM THE APOSTOLIC ADMINISTRATOR TOGETHER IN THE DIOCESE OF WAGGA WAGGA Eternal Rest Grant is published monthly, eleven times per year, and is available free at parish churches, schools, presbyteries and religious houses throughout the Wagga Wagga Unto them, O Lord Diocese. Archbishop Christopher Prowse - Apostolic Administrator of the Wagga Wagga Diocese Publisher The Diocese of Wagga Wagga Every year on 2nd November we poems and popular music. Too often the prepared a short document on”Guidelines for McAlroy House pray in a special way for those who richness of liturgical prayers and biblical Catholic Funerals.” It will be available on 205 Tarcutta Street have died before us. It is called The readings developed in the Catholic Church line in these days. Wagga Wagga NSW 2650 Commemoration of all the Faithful since ancient times can be put aside or even In this document there are some guidelines Editor/Designer Departed (All Souls Day). dismissed. for the place of eulogies, the place of Karene Eggleton Indeed, the whole of November is a special Thus, the preparation of funerals in the PowerPoint photo presentations, titles of the Printer Catholic Church is a pastoral challenge ceremonies, liturgical music, and cremations. month for us to pray for all who have The Riverina Media Group died. They are our “brothers and sisters” requiring great sensitivity from all of us. In regards to the latter, it needs to be noted Peter Street Wagga Wagga 2650 in the death and resurrection of Jesus. -
Murrumbidgee River EXPLORE • RESPECT • ENJOY • ESCAPE River Wisdom It Is Important to Enjoy the River Responsibly and Safely
EXPLORE • RESPECT • ENJOY • ESCAPE TO THE Murrumbidgee River EXPLORE • RESPECT • ENJOY • ESCAPE River Wisdom It is important to enjoy the river responsibly and safely. The following tips will help make the most of your visit and maintain the ecological integrity. Please contact the closest Visitors Information Centre for more information. Fishing The prime target species for anglers is the Murray Cod, followed by Golden Perch (Yellow Belly), Trout and Murray Crayfish. A NSW Recreational Fishing Fee (License) is required. There are fishing closures near weirs and fishing gear restrictions generally apply. Trout Cod, Macquarie Perch, Silver Perch, Freshwater Catfish and River Blackfish are totally protected. Be aware of the sizes, bag limits and seasons to ensure the continued protection of these species. Camping Visitors can camp on reserves but it is important to keep to areas marked as public land. It is advisable to take your own drinking water. Swimming/Water Sports Before swimming, boating or partaking in other water activities, care should be taken to test the depth, and the current of the river. Also be sure to check for submerged objects and take note of other river users. Children should always be supervised in and around water. Waste All visitors to the river are responsible for the removal of their own rubbish. Fruit Fly The Riverina is an important food producer and fruit fly can have a devastating impact on production. Please abide by the Fruit Fly Exclusion Zone signage and use roadside disposal bins. Lighting Fires There may be firebans at any time of the year. It is important you adhere to these.