Sorghum Malts for the Production of a Lager Beer
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Brewing Grains What Is Malt?
612.724.4514 [email protected] www.aperfectpint.net Brewing Grains Brewing grains are the heart and soul of beer. Next to water they make up the bulk of brewing ingredients. Brewing grains provide the sugars that yeast ferment. They are the primary source of beer color and a major contributor to beer flavor, aroma, and body. Proteins in the grains give structure to beer foam and minerals deliver many of the nutrients essential to yeast growth. By far the most common brewing grain is malted barley or barley malt, but a variety of other grains, both malted and unmalted, are also used including wheat, corn, rice, rye, and oats. What is Malt? To put it plainly, malt is cereal grain that has undergone the malting process. In the simplest terms, malting is the controlled germination and kilning of grain. Malting develops the diastatic enzymes that accomplish the conversion of starch to sugar during brewing and begins a limited process of conversion that makes the starches more accessible to the brewer. Malting also gives brewing grains their distinctive colors and flavors. Only the highest quality grain, called brewing grade, is selected for malting. Brewing grade grain is selected for, among other things, high starch content, uniform kernel size, low nitrogen content, and high diastatic power. Diastatic power is the ability of grains to break down complex starch molecules into simpler sugars for brewing. It is determined by the amount of diastatic enzymes in the grain. Barley is the most commonly malted grain, but other grains like wheat and rye are also malted. -

KAMUT® Brand Khorasan Wheat Whole Grain US Senator For
The Ancient Grain for Modern Life—Our mission is to promote organic agriculture and support organic farmers, to increase diversity of crops and diets and to protect the heritage of a high quality, delicious an- January 2013 cient grain for the benefit of this and future generations. Eat the Whole Thing: KAMUT® Brand Khorasan Wheat Whole Grain UPCOMING Whole grains are an important and tasty way of including complex carbohydrates in a healthy EVENTS diet. Depending on your age, health, weight, and activity level, the USDA recommends that Americans consume at least three portions, from 1.5 ounces (young children) to 8 ounces (older 20 – 22 January boys and young adult men) of grains a day, and that more than half of those grains should be 2013* - National As- whole grains. The US Food and Drug Administration (FDA) defines “whole grain to include cere- sociation for the Spe- al grains that consist of the intact, ground, cracked or flaked fruit of the grains whose principal cialty Food Trade, components -- the starchy endosperm, germ and bran -- are present in the same relative propor- Fancy Food Show, San tions as they exist in the intact grain.” Francisco, CA, USA Each part of the grain is healthful, but consuming them “whole” provides all of the benefits work- ing together. The FDA recognizes that whole grains provide energy and provide reduced risk for 25 January 2013 – disease including bowl disorders, cancer, heart disease and high cholesterol, stroke, high blood Annual KAMUT® Grower’s Dinner, Re- pressure, obesity and Type 2 diabetes. gina, SK, Canada In order to help you find good whole grain products, a lot of packaging includes the helpful term “whole grain” on the front or even better includes Whole Grains Council stamps. -
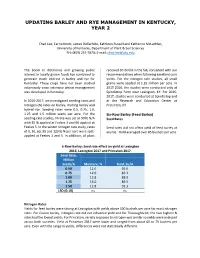
Updating Barley and Rye Management in Kentucky, Year 2
UPDATING BARLEY AND RYE MANAGEMENT IN KENTUCKY, YEAR 2 Chad Lee, Carrie Knott, James Dollarhide, Kathleen Russell and Katherine McLachlan, University of Kentucky, Department of Plant & Soil Sciences PH: (859) 257-7874; E-mail: [email protected] The boom in distilleries and growing public received 30 lb N/A in the fall, consistent with our interest in locally grown foods has combined to recommendations when following excellent corn generate much interest in barley and rye for yields. For the nitrogen rate studies, all small Kentucky. These crops have not been studied grains were seeded at 1.25 million per acre. In extensively since intensive wheat management 2015-2016, the studies were conducted only at was developed in Kentucky. Spindletop Farm near Lexington, KY. For 2016- 2017, studies were conducted at Spindletop and In 2016-2017, we investigated seeding rates and at the Research and Education Center at nitrogen (N) rates on barley, malting barley and Princeton, KY. hybrid rye. Seeding rates were 0.5, 0.75, 1.0, 1.25 and 1.5 million seeds per acre. For the Six-Row Barley (Feed Barley) seeding rate studies, N rate was set at 90 lb N/A Seed Rates with 30 lb applied at Feekes 3 and 60 applied at Feekes 5. In the winter nitrogen rate study, rates Seed rates did not affect yield of feed barley at of 0, 30, 60, 90 and 120 lb N per acre were split- any tie. Yield averaged over 85 bushels per acre. applied at Feekes 3 and 5. In addition, all plots 6-Row Barley: Seed rate effect on yield at Lexington 2016, Lexington 2017 and Princeton 2017. -

Coverage for Specialty Types of Barley and Soybeans Beginning with the 2010 Crop Year
United States May 27, 2010 Department of Agriculture INFORMATIONAL MEMORANDUM: PM-10-005.1 Risk Management TO: All Approved Insurance Providers Agency All Risk Management Agency Field Offices Beacon Facility – All Other Interested Parties Mail Stop 0812 P.O. Box 419205 FROM: Tim B. Witt /s/ Tim B. Witt Kansas City, MO 64141-6205 Deputy Administrator SUBJECT: Coverage for Specialty Types of Barley and Soybeans Beginning with the 2010 Crop Year BACKGROUND: On February 1, 2010, the Risk Management Agency (RMA) issued PM-10-005 regarding insurance coverage for specialty types of barley and soybeans based on contract prices effective for the 2010 crop year. Some questions have arisen pertaining to type identification, prevented planting, replanting, and quality adjustment. RMA is providing the following guidance when insuring specialty types. ACTION: 1. Identifying Specialty Types: There are six possible types specified in the Special Provisions of Insurance (SPOI): All Others (AO), and the five specialty types shown below with the requirements for each: a. Large seeded food grade – Soybeans commonly used for tofu, soymilk, and miso having a minimum seed size of 20g/100 seeds. b. Small seeded food grade – Soybeans commonly used for sprouts having a minimum seed size of 10g/100 seeds, or for natto soybeans having a minimum seed size of 8g/100 seeds. c. Low linolenic acid – Soybeans commonly used to produce soybean oil with a linolenic acid level of three percent or less. d. Low saturated fat - Soybeans containing 50 percent less saturated fat than conventional soybeans and are used to produce soybean oil with eight percent or less total saturated fats. -
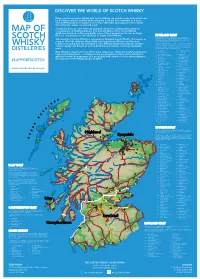
2019 Scotch Whisky
©2019 scotch whisky association DISCOVER THE WORLD OF SCOTCH WHISKY Many countries produce whisky, but Scotch Whisky can only be made in Scotland and by definition must be distilled and matured in Scotland for a minimum of 3 years. Scotch Whisky has been made for more than 500 years and uses just a few natural raw materials - water, cereals and yeast. Scotland is home to over 130 malt and grain distilleries, making it the greatest MAP OF concentration of whisky producers in the world. Many of the Scotch Whisky distilleries featured on this map bottle some of their production for sale as Single Malt (i.e. the product of one distillery) or Single Grain Whisky. HIGHLAND MALT The Highland region is geographically the largest Scotch Whisky SCOTCH producing region. The rugged landscape, changeable climate and, in The majority of Scotch Whisky is consumed as Blended Scotch Whisky. This means as some cases, coastal locations are reflected in the character of its many as 60 of the different Single Malt and Single Grain Whiskies are blended whiskies, which embrace wide variations. As a group, Highland whiskies are rounded, robust and dry in character together, ensuring that the individual Scotch Whiskies harmonise with one another with a hint of smokiness/peatiness. Those near the sea carry a salty WHISKY and the quality and flavour of each individual blend remains consistent down the tang; in the far north the whiskies are notably heathery and slightly spicy in character; while in the more sheltered east and middle of the DISTILLERIES years. region, the whiskies have a more fruity character. -

The Whiskey Machine: Nanofactory-Based Replication of Fine Spirits and Other Alcohol-Based Beverages
The Whiskey Machine: Nanofactory-Based Replication of Fine Spirits and Other Alcohol-Based Beverages © 2016 Robert A. Freitas Jr. All Rights Reserved. Abstract. Specialized nanofactories will be able to manufacture specific products or classes of products very efficiently and inexpensively. This paper is the first serious scaling study of a nanofactory designed for the manufacture of a specific food product, in this case high-value-per- liter alcoholic beverages. The analysis indicates that a 6-kg desktop appliance called the Fine Spirits Synthesizer, aka. the “Whiskey Machine,” consuming 300 W of power for all atomically precise mechanosynthesis operations, along with a commercially available 59-kg 900 W cryogenic refrigerator, could produce one 750 ml bottle per hour of any fine spirit beverage for which the molecular recipe is precisely known at a manufacturing cost of about $0.36 per bottle, assuming no reduction in the current $0.07/kWh cost for industrial electricity. The appliance’s carbon footprint is a minuscule 0.3 gm CO2 emitted per bottle, more than 1000 times smaller than the 460 gm CO2 per bottle carbon footprint of conventional distillery operations today. The same desktop appliance can intake a tiny physical sample of any fine spirit beverage and produce a complete molecular recipe for that product in ~17 minutes of run time, consuming <25 W of power, at negligible additional cost. Cite as: Robert A. Freitas Jr., “The Whiskey Machine: Nanofactory-Based Replication of Fine Spirits and Other Alcohol-Based Beverages,” IMM Report No. 47, May 2016; http://www.imm.org/Reports/rep047.pdf. 2 Table of Contents 1. -

How to Malt Barley (Or Wheat) for Beer (Or Whisky/Whiskey) I
How to Malt Barley (or Wheat) for Beer (or Whisky/Whiskey) I. Start with Good Quality Barley (or Wheat) A. When purchasing grain for malting, first ask about what types of chemicals might have been used on the grain and then about the grain's history: when it was harvested, how it was stored, etc. B. Inspect the Grain 1. Bits of chaff – and even the occasional dead bug – are quite normal and are of little concern. This sort of debris can easily be removed later when the grain is washed. It will usually float to the surface of the water where it can be skimmed off in a matter of minutes. 2. Grain that has live bugs should generally be avoided, as such bugs will spread to other grain stores and feast on all available grain until it is ready for use. Some of those bugs will also contribute a strong off-smell and off-taste, as well. However, buggy grain can occasionally be had for free, and there are a few types of bugs that do minimal damage to the grain if they are not allowed much time to do their worst. But definitely proceed cautiously with live bugs. 3. The presence of foreign seeds mixed in with the grain may or may not be problematic. A few kernels of corn, a bean or two, a few oats, etc., should not be cause for concern, but if there are unknown seeds mixed in with the grain, ask what they are and act accordingly. 4. Examine the color of the grain and avoid any that has a grayish hue, as this would indicate that the grain was mildewed, either in the field or in storage. -

Undestanding Malting Barley Quality Aaron Macleod, Director, Center for Craft Food and Beverage Hartwick College, Oneonta, NY, 13820 [email protected]
Undestanding Malting Barley Quality Aaron MacLeod, Director, Center for Craft Food and Beverage Hartwick College, Oneonta, NY, 13820 [email protected] Malt is the key ingredient in beer that provides the starch and enzymes necessary to produce the fermentable sugars which yeast then turn into alcohol. Malt also provides the color and flavor compounds which contribute to the final character of beer. Malting is the biological process that turns barley into malt. It is a three stage process including soaking (or steeping) the grain in water to bring the kernels to 45% moisture; germination under cool, humid conditions and drying (or kilning) to dry and stabilize the final malt. Barley must meet strict quality criteria to be acceptable for malt production. Maintaining tight controls on these quality factors in the grain is necessary to ensure good processing efficiency and final product quality in the malthouse and brewery. Care must be taken when growing malting barley to ensure that it meets the necessary specifications. A premium is paid to growers for high quality malting barley to compensate for the extra effort. High quality malting barley should have the following characteristics: Pure lot of an acceptable variety Germination of 95% or higher Protein content raging between 9.5% to 12.5% (dry basis) Moisture content below 13.5% Plump and uniform kernels Free of disease and low DON content Less than 5% of peeled, broken, or damaged kernels Clean and free of insects, admixtures, ergot or foreign material Varietal purity Malting barley varieties are bred specifically for characteristics that promote good malting and brewing performance, such as high enzymatic activity, as well as good agronomic performance and disease resistance. -

Wheat, Barley, Rye, GO! Students Get Active and Learn About Whole Grains in This Spirited Game Overview
Wheat, Barley, Rye, GO! Students get active and learn about whole grains in this spirited game Overview In this wacky version of "Rock, Paper, Scissors," students strategize and chase Description each other while learning about whole grains. Objective Students will identify a variety of whole grain foods they can eat for snack. Activity 1. Have the studends form a large circle. 2. Ask them to raise their hands if they eat whole grain foods for snack (e.g. crackers, bread, etc). Explain that grains are carbohydrates, the body’s main source of energy. Tell the class that whole grains are usually brown and are healthier than white grains because they have more vitamins and nutrients, which give the body more energy to run and play. 3. Then, ask them to share a few specific whole grain foods they eat. (If a student mentions a processed, "white" grain such as white bread, tell them it is okay to eat foods like white bread once in a while, but they should eat whole grains more often. Can they think of a whole grain food to replace the other?) 4. Divide the class into two groups and have them stand at opposite ends of the room. 5. Explain that they are going to play a familiar game "Rock, Paper, Scissors" with a twist. The name of the game is "Wheat, Barley, Rye." 6. Have the class create one full-body pose (as opposed to hand sign) for each grain. Have the students practice the movements as you call out the grains so they become familiar. -

Breakdown of a Malt COA
Breakdown of a Malt COA Bucket Analysis Approach Presenters Tyler Schoales Mike Heinrich Craft Malt Specialist – NA Craft Sales Manager Country Malt Group Great Western Malting Breakdown of a Malt COA Agenda Overview of Malting and Modification Certificate of Analysis Breakdown Bucketing Analysis • Protein Dependent Specifications • Carbohydrate Dependent Specifications • Enzyme Package Specifications • Color Breakdown of a Malt COA Malting and Breakdown of Bucketing Modification COA Analysis Malting and the Certificate of Analysis • Malting is the controlled germination and kilning of a seed to produce the desirable brewing characteristics • Maltsters create ideal growing conditions for barley to germinate and drive modification • “Modification” is the biochemical breakdown of cell wall structures, and protein matrices in order to gain access to the starch reserves held within the endosperm • A malt Certificate of Analysis (COA) lists the results from a suite of standardized tests that serve to indicate how the malt will perform. Breakdown of a Malt COA Malting and Breakdown of Bucketing Modification COA Analysis Breakdown of the COA Malt Sieve Analysis (Assortment) • Plump kernels provide more extract than thinner kernels • Roller mill gaps are set according to the mean kernel size • A broad distribution can make mill setting difficult → poor extract recovery in the brewery • Typical analysis – 7/64 + 6/64’s (PLUMP’s) > 90% • Consistency is the key Malt Sieve Analysis (Breakage) • Damaged husks will form a poor filter bed • Fines formed -

FARM Energy Success Stories
New Farm INcome Farm energy Success Stories New Farm INcome · rur al ecoNomIc DevelopmeNt Energy IndependeNce · a CLEANER EnvIroNmeNt CongReSSIonaL SuppoRT foR THe faRm BILL’S CLean eneRgy pRogRamS “American farmers and rural businesses “Our insatiable appetite for energy, particularly are successfully using the Rural Energy for from outside our borders, represents one of our America Program to leverage billions of gravest security threats. The Farm Bill’s energy dollars in private investment for successful programs recognize our nation’s agriculture new renewable energy and energy efficiency and rural sector’s ability to confront these risks. projects. These new energy projects are good Solutions such as REAP improve our nation’s energy for rural economies, good for the environment equation and help strengthen our rural economies.” and good for our national energy security.” —Senator Tom Harkin (D-Iowa) —Senator Richard Lugar (R-Indiana) “Rural America possesses the resources and spirit “I strongly support the Rural Energy for America to lead our nation away from dependence on Program because it is one of the only federal programs foreign oil and non-renewable sources of energy. that comprehensively transforms a clean energy Building on the 2002 Farm Bill efforts, the development vision into action across agricultural 2008 Farm Bill is now expanding opportunities America. From the time when we created this for farmers, ranchers and small businesses to program in the 2002 Farm Bill to the nationally conserve energy and produce clean renewable successful program today, we have cemented our energy. Programs like the Rural Energy for commitment to transition rural and agricultural America Program provide the resources to America toward a future that treats energy as a transform practical ideas to save or produce strategic resource which must be conserved, protected energy into a reality. -

Consumer Demand for Simple, Healthy and Nutritious Foods Leads to Rise of Ancient Grains
PR Contact: Jillian Chertok, ADinfinitum, New York, NY. Email: [email protected] Tel: 212.693.2150 Ext: 311 FOR IMMEDIATE RELEASE Consumer Demand for Simple, Healthy and Nutritious Foods Leads to Rise of Ancient Grains Sales of ancient grains such as KAMUT® Brand khorasan wheat, quinoa, amaranth, teff, spelt and freekah have increased dramatically in recent years as more consumers, nutritionists and big brands recognize their benefits Big Sandy, MT, September 2015 – Consumers and nutrition experts have spoken – driven by their desire for non-GMO foods with simple ingredients that are high in nutrients, ancient grains are on the rise with no sign of slowing down. In fact, according to data from SPINS, a leading supplier of retail consumer analytics and insights, sales of ancient grains rose steeply in the 52 weeks ending July 13, 2014. All ancient grains, such as KAMUT®, amaranth, quinoa, spelt and freekah, will continue to grow through 2015 and will be a top nutritional trend, according to a recent survey of more than 500 registered dietitians from Today’s Dietitian and Pollock Communications. Unlike modern grains, ancient grains have survived intact for centuries and have more vitamins, minerals, fiber and proteins than modern grains, as well as more distinctive and flavorful taste. Due to their rise in popularity and demand, ancient grains are now more accessible than ever before, especially as well known brands, such as Kashi® and Kellogg’s, have started to incorporate them into their products. In August 2015, Kashi® released its Organic Promise® Cocoa Coconut with KAMUT® Khorasan Wheat Granola, which pairs ancient grains with superfoods, including oats and coconut.