Hymenoptera: Apidae Sl: Protandrenini
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Chemical Ecology and Evolution of Bee–Flower Interactions: a Review and Perspectives1
668 REVIEW / SYNTHE` SE The chemical ecology and evolution of bee–flower interactions: a review and perspectives1 S. Do¨ tterl and N.J. Vereecken Abstract: Bees and angiosperms have shared a long and intertwined evolutionary history and their interactions have re- sulted in remarkable adaptations. Yet, at a time when the ‘‘pollination crisis’’ is of major concern as natural populations of both wild and honey bees (Apis mellifera L., 1758) face alarming decline rates at a worldwide scale, there are important gaps in our understanding of the ecology and evolution of bee–flower interactions. In this review, we summarize and dis- cuss the current knowledge about the role of floral chemistry versus other communication channels in bee-pollinated flow- ering plants, both at the macro- and micro-evolutionary levels, and across the specialization–generalization gradient. The available data illustrate that floral scents and floral chemistry have been largely overlooked in bee–flower interactions, and that pollination studies integrating these components along with pollinator behaviour in a phylogenetic context will help gain considerable insights into the sensory ecology and the evolution of bees and their associated flowering plants. Re´sume´ : Les abeilles et les angiospermes partagent une grande partie de leur histoire e´volutive, et leurs interactions ont produit de remarquables exemples d’adaptations mutuelles. Cependant, a` une e´poque ou` la « crise de la pollinisation » de- vient une pre´occupation majeure et ou` les populations d’abeilles sauvages et mellife`res (Apis mellifera L., 1758) font face a` des de´clins massifs a` l’e´chelle mondiale, notre compre´hension de l’e´cologie et de l’e´volution des relations abeilles- plantes demeure fragmentaire. -

Solanaceae) Flower–Visitor Network in an Atlantic Forest Fragment in Southern Brazil
diversity Article Bee Diversity and Solanum didymum (Solanaceae) Flower–Visitor Network in an Atlantic Forest Fragment in Southern Brazil Francieli Lando 1 ID , Priscila R. Lustosa 1, Cyntia F. P. da Luz 2 ID and Maria Luisa T. Buschini 1,* 1 Programa de Pós Graduação em Biologia Evolutiva da Universidade Estadual do Centro-Oeste, Rua Simeão Camargo Varela de Sá 03, Vila Carli, Guarapuava 85040-080, Brazil; [email protected] (F.L.); [email protected] (P.R.L.) 2 Research Centre of Vascular Plants, Palinology Research Centre, Botanical Institute of Sao Paulo Government, Av. Miguel Stéfano, 3687 Água Funda, São Paulo 04045-972, Brazil; [email protected] * Correspondence: [email protected] Received: 9 November 2017; Accepted: 8 January 2018; Published: 11 January 2018 Abstract: Brazil’s Atlantic Forest biome is currently undergoing forest loss due to repeated episodes of devastation. In this biome, bees perform the most frequent pollination system. Over the last decade, network analysis has been extensively applied to the study of plant–pollinator interactions, as it provides a consistent view of the structure of plant–pollinator interactions. The aim of this study was to use palynological studies to obtain an understanding of the relationship between floral visitor bees and the pioneer plant S. didymum in a fragment of the Atlantic Forest, and also learn about the other plants that interact to form this network. Five hundred bees were collected from 32 species distributed into five families: Andrenidae, Apidae, Colletidae, Megachilidae, and Halictidae. The interaction network consisted of 21 bee species and 35 pollen types. -

Apoidea: Andrenidae: Panurginae)
Contemporary distributions of Panurginus species and subspecies in Europe (Apoidea: Andrenidae: Panurginae) Sébastien Patiny Abstract The largest number of Old World Panurginus Nylander, 1848 species is distributed in the West- Palaearctic. The genus is absent in Africa and rather rare in the East-Palaearctic. Warncke (1972, 1987), who is the main author treating Palaearctic Panurginae in the last decades, subdivided Panurginus into a small number of species, including two principal taxa admitting for each a very large number of subspecies: Panurginus brullei (Lepeletier, 1841) and Panurginus montanus Giraud, 1861. Following recent works, the two latter are in fact complexes of closely related species. In the West-Palaearctic context, distributions of certain species implied in these complexes appear as very singular, distinct of mostly all other Panurginae ranges and highly interesting from a fundamental point of view. Based on a cartographic approach, the causes which influence (or have conditioned, in the past) the observed ranges of these species are discussed. Key words: Panurginae, biogeography, West-Palaearctic, speciation, glaciation. Introduction 1841) sensu lato are studied in the present paper. In the Old World Panurginae fauna, Panurginus The numerous particularities of their distribu- Nylander, 1848 is one of the only two genera tions are characterized and discussed here. The (with Melitturga Latreille, 1809) which are limits of these distributions were proposed and distributed in the entire Palaearctic region. The related to the contemporary and past develop- genera Camptopoeum Spinola, 1843 and Panur- ments which could have caused these ranges. gus Panzer, 1806 are also represented in the East- Palaearctic (in northern Thailand), but too few Catalogue of the old world species data are available for these genera to make them of Panurginus the subject of particular considerations. -
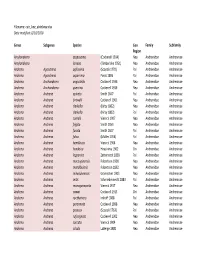
Anthophila List
Filename: cuic_bee_database. -

1. Padil Species Factsheet Scientific Name: Common Name Image
1. PaDIL Species Factsheet Scientific Name: Panurginus ineptus Cockerell, 1922 (Hymenoptera: Andrenidae: Panurginae: Panurgini) Common Name Tribe Representative - Panurgini Live link: http://www.padil.gov.au/pollinators/Pest/Main/139784 Image Library Australian Pollinators Live link: http://www.padil.gov.au/pollinators/ Partners for Australian Pollinators image library Western Australian Museum https://museum.wa.gov.au/ South Australian Museum https://www.samuseum.sa.gov.au/ Australian Museum https://australian.museum/ Museums Victoria https://museumsvictoria.com.au/ 2. Species Information 2.1. Details Specimen Contact: Museum Victoria - [email protected] Author: Ken Walker Citation: Ken Walker (2010) Tribe Representative - Panurgini(Panurginus ineptus)Updated on 8/11/2010 Available online: PaDIL - http://www.padil.gov.au Image Use: Free for use under the Creative Commons Attribution-NonCommercial 4.0 International (CC BY- NC 4.0) 2.2. URL Live link: http://www.padil.gov.au/pollinators/Pest/Main/139784 2.3. Facets Bio-Region: USA and Canada, Europe and Northern Asia Host Family: Not recorded Host Genera: Fresh Flowers Status: Exotic Species not in Australia Bio-Regions: Palaearctic, Nearctic Body Hair and Scopal location: Body hair relatively sparse, Tibia Episternal groove: Present but not extending below scrobal groove Wings: Submarginal cells - Three, Apex of marginal cell truncate or rounded Head - Structures: Two subantennal sutures below each antennal socket, Facial fovea present usually as a broad groove, Facial -

Hymenoptera, Apoidea)
>lhetian JMfuseum ox4tates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 2 24 AUGUST I7, I 965 The Biology and Immature Stages of Melitturga clavicornis (Latreille) and of Sphecodes albilabris (Kirby) and the Recognition of the Oxaeidae at the Family Level (Hymenoptera, Apoidea) BYJEROME G. ROZEN, JR.' Michener (1944) divided the andrenid subfamily Panurginae into two tribes, the Panurgini and Melitturgini, with the latter containing the single Old World genus Melitturga. This genus was relegated to tribal status apparently on the grounds that the adults, unlike those of other panurgines, bear certain striking resemblances to the essentially Neo- tropical Oxaeinae of the same family. In 1951 Rozen showed that the male genitalia of Melitturga are unlike those of the Oxaeinae and are not only typical of those of the Panurginae in general but quite like those of the Camptopoeum-Panurgus-Panurginus-Epimethea complex within the sub- family. On the basis of this information, Michener (1954a) abandoned the idea that the genus Melitturga represents a distinct tribe of the Panur- ginae. Recently evidence in the form of the larva of Protoxaea gloriosa Fox (Rozen, 1965) suggested that the Oxaeinae were so unlike other Andreni- dae that they should be removed from the family unless some form inter- 1 Chairman and Associate Curator, Department of Entomology, the American Museum of Natural History. 2 AMERICAN MUSEUM NOVITATES NO. 2224 mediate between the two subfamilies is found. In spite of the structure of the male genitalia, Melitturga is the only known possible intermediary. -

Novitates PUBLISHED by the AMERICAN MUSEUM of NATURAL HISTORY CENTRAL PARK WEST at 79TH STREET, NEW YORK, N.Y
AMERICAN MUSEUM Novitates PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK, N.Y. 10024 Number 2640, pp. 1-24, figs. 1-36, tables 1-3 January 3, 1978 The Bionomics and Immature Stages of the Cleptoparasitic Bee Genus Protepeolus (Anthophoridae, Nomadinae) JEROME G. ROZEN, JR.,' KATHLEEN R. EICKWORT,2 AND GEORGE C. EICKWORT3 ABSTRACT Protepeolus singularis was found attacking cells numerous biological dissimilarities. The first in- in nests of Diadasia olivacea in southeastern Ari- star Protepeolus attacks and kills the pharate last zona. The following biological information is pre- larval instar of the host before consuming the sented: behavior of adult females while searching provisions, a unique feature for nomadine bees. for host nests; intraspecific interactions of fe- First and last larval instars and the pupa are males at the host nesting site; interactions with described taxonomically and illustrated. Brief host adults; oviposition; and such larval activities comparative descriptions of the other larval in- as crawling, killing the host, feeding, defecation, stars are also given. Larval features attest to the and cocoon spinning. In general, adult female be- common origin of Protepeolus and the other havior corresponds to that of other Nomadinae. Nomadinae. Cladistic analysis using 27 characters Females perch for extended periods near nest of mature larvae of the Nomadinae demonstrates entrances and avoid host females, which attack that Isepeolus is a sister group to all the other parasites when encountered. Females apparently Nomadinae known from larvae, including Pro- learn the locations of host nests and return to tepeolus, and that Protepeolus is a sister group to them frequently. -

Journal of Hymenoptera Research
c 3 Journal of Hymenoptera Research . .IV 6«** Volume 15, Number 2 October 2006 ISSN #1070-9428 CONTENTS BELOKOBYLSKIJ, S. A. and K. MAETO. A new species of the genus Parachremylus Granger (Hymenoptera: Braconidae), a parasitoid of Conopomorpha lychee pests (Lepidoptera: Gracillariidae) in Thailand 181 GIBSON, G. A. P., M. W. GATES, and G. D. BUNTIN. Parasitoids (Hymenoptera: Chalcidoidea) of the cabbage seedpod weevil (Coleoptera: Curculionidae) in Georgia, USA 187 V. Forest GILES, and J. S. ASCHER. A survey of the bees of the Black Rock Preserve, New York (Hymenoptera: Apoidea) 208 GUMOVSKY, A. V. The biology and morphology of Entedon sylvestris (Hymenoptera: Eulophidae), a larval endoparasitoid of Ceutorhynchus sisymbrii (Coleoptera: Curculionidae) 232 of KULA, R. R., G. ZOLNEROWICH, and C. J. FERGUSON. Phylogenetic analysis Chaenusa sensu lato (Hymenoptera: Braconidae) using mitochondrial NADH 1 dehydrogenase gene sequences 251 QUINTERO A., D. and R. A. CAMBRA T The genus Allotilla Schuster (Hymenoptera: Mutilli- dae): phylogenetic analysis of its relationships, first description of the female and new distribution records 270 RIZZO, M. C. and B. MASSA. Parasitism and sex ratio of the bedeguar gall wasp Diplolqjis 277 rosae (L.) (Hymenoptera: Cynipidae) in Sicily (Italy) VILHELMSEN, L. and L. KROGMANN. Skeletal anatomy of the mesosoma of Palaeomymar anomalum (Blood & Kryger, 1922) (Hymenoptera: Mymarommatidae) 290 WHARTON, R. A. The species of Stenmulopius Fischer (Hymenoptera: Braconidae, Opiinae) and the braconid sternaulus 316 (Continued on back cover) INTERNATIONAL SOCIETY OF HYMENOPTERISTS Organized 1982; Incorporated 1991 OFFICERS FOR 2006 Michael E. Schauff, President James Woolley, President-Elect Michael W. Gates, Secretary Justin O. Schmidt, Treasurer Gavin R. -

The Very Handy Bee Manual
The Very Handy Manual: How to Catch and Identify Bees and Manage a Collection A Collective and Ongoing Effort by Those Who Love to Study Bees in North America Last Revised: October, 2010 This manual is a compilation of the wisdom and experience of many individuals, some of whom are directly acknowledged here and others not. We thank all of you. The bulk of the text was compiled by Sam Droege at the USGS Native Bee Inventory and Monitoring Lab over several years from 2004-2008. We regularly update the manual with new information, so, if you have a new technique, some additional ideas for sections, corrections or additions, we would like to hear from you. Please email those to Sam Droege ([email protected]). You can also email Sam if you are interested in joining the group’s discussion group on bee monitoring and identification. Many thanks to Dave and Janice Green, Tracy Zarrillo, and Liz Sellers for their many hours of editing this manual. "They've got this steamroller going, and they won't stop until there's nobody fishing. What are they going to do then, save some bees?" - Mike Russo (Massachusetts fisherman who has fished cod for 18 years, on environmentalists)-Provided by Matthew Shepherd Contents Where to Find Bees ...................................................................................................................................... 2 Nets ............................................................................................................................................................. 2 Netting Technique ...................................................................................................................................... -

Along Urbanization Sprawl, Exotic Plants Distort Native Bee (Hymenoptera: Apoidea) Assemblages in High Elevation Andes Ecosystem
A peer-reviewed version of this preprint was published in PeerJ on 7 November 2018. View the peer-reviewed version (peerj.com/articles/5916), which is the preferred citable publication unless you specifically need to cite this preprint. Henríquez-Piskulich P, Vera A, Sandoval G, Villagra C. 2018. Along urbanization sprawl, exotic plants distort native bee (Hymenoptera: Apoidea) assemblages in high elevation Andes ecosystem. PeerJ 6:e5916 https://doi.org/10.7717/peerj.5916 Along urbanization sprawl, exotic plants distort native bee (Hymenoptera: Apoidea) assemblages in high elevation Andes ecosystem Patricia Henríquez-Piskulich Corresp., 1 , Alejandro Vera 2 , Gino Sandoval 3 , Cristian Villagra 1 1 Instituto de Entomología, Universidad Metropolitana de Ciencias de la Educación, Santiago, Región Metropolitana, Chile 2 Departamento de Biología, Universidad Metropolitana de Ciencias de la Educación, Santiago, Región Metropolitana, Chile 3 Departamento de Historia y Geografía, Universidad Metropolitana de Ciencias de la Educación, Santiago, Región Metropolitana, Chile Corresponding Author: Patricia Henríquez-Piskulich Email address: [email protected] Native bees contribute with a considerable portion of pollination services for endemic as well as economically important plant species. Their decline has been attributed to several human-derived influences including global warming as well as the reduction, alteration and loss of bees’ habitat. Moreover, together with human expansion comes along the introduction of exotic plant species with negative impacts over native ecosystems. Anthropic effects may have even a deeper impact on communities adapted to extreme environments, such as high elevation habitats, where abiotic stressors alone are a natural limitation to biodiversity. In these, human-borne alterations, such as the introduction of exotic plants and urbanization, may have a greater influence on native communities. -

A Catalogue of the Family Andrenidae (Hymenoptera: Apoidea) of Eritrea
Linzer biol. Beitr. 50/1 655-659 27.7.2018 A catalogue of the family Andrenidae (Hymenoptera: Apoidea) of Eritrea Michael MADL A b s t r a c t : In Eritrea the family Andrenidae is represented by two species of the genus Andrena FABRICIUS, 1775 (subfamily Andreninae) and one species each oft he genera Borgatomelissa PATINY, 2000 and Meliturgula FRIESE, 1903 (both subfamily Panurginae). The record of the genus Andrena is doubtful. K e y w o r d s : Andrenidae, Andreninae, Andrena, Panurginae, Borgatomelissa, Meliturgula, Eritrea. Introduction In the Afrotropical region the family Andrenidae is represented by more than 30 species (EARDLEY & URBAN 2010). The Eritrean fauna of of the family Andrenidae is poorly known. The knowledge is based on the material collected by J.K. Lord in 1869 and by P. Magretti in 1900. No further material is known. Up to date two species each of the subfamilies Andreninae (genus Andrena FABRICIUS, 1775) and Panurginae (genera Borgatomelissa PATINY, 2000 and Meliturgula FRIESE, 1903) have been recorded from Eritrea. Abbreviations Afr. reg. .................................. Afrotropical region biogeogr. ......................................... biogeography biol. .......................................................... biology cat. ......................................................... catalogue descr. ................................................... description fig. (figs) ....................................... figure (figures) syn. ......................................................... synonym tab. -

American Museum Novitates
AMERICAN MUSEUM NOVITATES Number 3814, 16 pp. October 16, 2014 Nesting biology and immature stages of the panurgine bee genera Rhophitulus and Cephalurgus (Apoidea: Andrenidae: Protandrenini) JEROME G. ROZEN, JR.1 ABSTRACT Herein is presented nesting information on the communal ground-nesting Argentinian bees Rhophitulus xenopalpus Ramos and R. mimus Ramos, which is compared with what is known concerning the closely related Brazilian bee Cephalurgus anomalus Moure and Lucas de Oliveira. The mature larvae of all three taxa are described, illustrated, and compared with one another and with those of other Protandrenini. While larvae of the three species share many similarities, those of R. xenopalpus and R. mimus, though each distinctive, are quite similar, and those of C. anomalus differ from the others in mandibular features and in dorsal body orna- mentation. Male and female pupae of R. xenopalpus are also described. INTRODUCTION The purpose of this project was to advance our understanding of the nesting biology and immature stages of bees belonging to the panurgine genera Rhophitulus and Cephalurgus, a complex of small, obscure, mostly black, ground-nesting species restricted to South America (Michener, 2007). Presented in this report is information on the nesting biology of Rhophitulus xenopalpus Ramos and R. mimus Ramos, which is compared with the previously published biological account of Cephalurgus anomalus Moure and Lucas de Oliveira (Rozen, 1989). Although in the past consid- ered only subgenerically distinct (Michener, 2007), Rhophitulus and Cephalurgus are now thought to be separate genera (Moure et al., 2007, 2012). Accordingly, their separate relationship is main- tained here, and the two genera are treated together to be further compared.