Interdependence of Torque, Joint Angle, Angular Velocity and Muscle Action During Human Multi-Joint Leg Extension
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rotational Motion (The Dynamics of a Rigid Body)
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Robert Katz Publications Research Papers in Physics and Astronomy 1-1958 Physics, Chapter 11: Rotational Motion (The Dynamics of a Rigid Body) Henry Semat City College of New York Robert Katz University of Nebraska-Lincoln, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/physicskatz Part of the Physics Commons Semat, Henry and Katz, Robert, "Physics, Chapter 11: Rotational Motion (The Dynamics of a Rigid Body)" (1958). Robert Katz Publications. 141. https://digitalcommons.unl.edu/physicskatz/141 This Article is brought to you for free and open access by the Research Papers in Physics and Astronomy at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Robert Katz Publications by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. 11 Rotational Motion (The Dynamics of a Rigid Body) 11-1 Motion about a Fixed Axis The motion of the flywheel of an engine and of a pulley on its axle are examples of an important type of motion of a rigid body, that of the motion of rotation about a fixed axis. Consider the motion of a uniform disk rotat ing about a fixed axis passing through its center of gravity C perpendicular to the face of the disk, as shown in Figure 11-1. The motion of this disk may be de scribed in terms of the motions of each of its individual particles, but a better way to describe the motion is in terms of the angle through which the disk rotates. -

Velocity-Corrected Area Calculation SCIEX PA 800 Plus Empower
Velocity-corrected area calculation: SCIEX PA 800 Plus Empower Driver version 1.3 vs. 32 Karat™ Software Firdous Farooqui1, Peter Holper1, Steve Questa1, John D. Walsh2, Handy Yowanto1 1SCIEX, Brea, CA 2Waters Corporation, Milford, MA Since the introduction of commercial capillary electrophoresis (CE) systems over 30 years ago, it has been important to not always use conventional “chromatography thinking” when using CE. This is especially true when processing data, as there are some key differences between electrophoretic and chromatographic data. For instance, in most capillary electrophoresis separations, peak area is not only a function of sample load, but also of an analyte’s velocity past the detection window. In this case, early migrating peaks move past the detection window faster than later migrating peaks. This creates a peak area bias, as any relative difference in an analyte’s migration velocity will lead to an error in peak area determination and relative peak area percent. To help minimize Figure 1: The PA 800 Plus Pharmaceutical Analysis System. this bias, peak areas are normalized by migration velocity. The resulting parameter is commonly referred to as corrected peak The capillary temperature was maintained at 25°C in all area or velocity corrected area. separations. The voltage was applied using reverse polarity. This technical note provides a comparison of velocity corrected The following methods were used with the SCIEX PA 800 Plus area calculations using 32 Karat™ and Empower software. For Empower™ Driver v1.3: both, standard processing methods without manual integration were used to process each result. For 32 Karat™ software, IgG_HR_Conditioning: conditions the capillary Caesar integration1 was turned off. -

The Origins of Velocity Functions
The Origins of Velocity Functions Thomas M. Humphrey ike any practical, policy-oriented discipline, monetary economics em- ploys useful concepts long after their prototypes and originators are L forgotten. A case in point is the notion of a velocity function relating money’s rate of turnover to its independent determining variables. Most economists recognize Milton Friedman’s influential 1956 version of the function. Written v = Y/M = v(rb, re,1/PdP/dt, w, Y/P, u), it expresses in- come velocity as a function of bond interest rates, equity yields, expected inflation, wealth, real income, and a catch-all taste-and-technology variable that captures the impact of a myriad of influences on velocity, including degree of monetization, spread of banking, proliferation of money substitutes, devel- opment of cash management practices, confidence in the future stability of the economy and the like. Many also are aware of Irving Fisher’s 1911 transactions velocity func- tion, although few realize that it incorporates most of the same variables as Friedman’s.1 On velocity’s interest rate determinant, Fisher writes: “Each per- son regulates his turnover” to avoid “waste of interest” (1963, p. 152). When rates rise, cashholders “will avoid carrying too much” money thus prompting a rise in velocity. On expected inflation, he says: “When...depreciation is anticipated, there is a tendency among owners of money to spend it speedily . the result being to raise prices by increasing the velocity of circulation” (p. 263). And on real income: “The rich have a higher rate of turnover than the poor. They spend money faster, not only absolutely but relatively to the money they keep on hand. -

VELOCITY from Our Establishment in 1957, We Have Become One of the Oldest Exclusive Manufacturers of Commercial Flooring in the United States
VELOCITY From our establishment in 1957, we have become one of the oldest exclusive manufacturers of commercial flooring in the United States. As one of the largest privately held mills, our FAMILY-OWNERSHIP provides a heritage of proven performance and expansive industry knowledge. Most importantly, our focus has always been on people... ensuring them that our products deliver the highest levels of BEAUTY, PERFORMANCE and DEPENDABILITY. (cover) Velocity Move, quarter turn. (right) Velocity Move with Pop Rojo and Azul, quarter turn. VELOCITY 3 velocity 1814 style 1814 style 1814 style 1814 color 1603 color 1604 color 1605 position direction magnitude style 1814 style 1814 style 1814 color 1607 color 1608 color 1609 reaction move constant style 1814 color 1610 vector Velocity Vector, quarter turn. VELOCITY 5 where to use kinetex Healthcare Fitness Centers kinetex overview Acute care hospitals, medical Health Clubs/Gyms office buildings, urgent care • Cardio Centers clinics, outpatient surgery • Stationary Weight Centers centers, outpatient physical • Dry Locker Room Areas therapy/rehab centers, • Snack Bars outpatient imaging centers, etc. • Offices Kinetex® is an advanced textile composite flooring that combines key attributes of • Cafeteria, dining areas soft-surface floor covering with the long-wearing performance characteristics of • Chapel Retail / Mercantile hard-surface flooring. Created as a unique floor covering alternative to hard-surface Wholesale / Retail merchants • Computer room products, J+J Flooring’s Kinetex encompasses an unprecedented range of • Corridors • Checkout / cash wrap performance attributes for retail, healthcare, education and institutional environments. • Diagnostic imaging suites • Dressing rooms In addition to its human-centered qualities and highly functional design, Kinetex • Dry physical therapy • Sales floor offers a reduced environmental footprint compared to traditional hard-surface options. -

Chapter 5 the Relativistic Point Particle
Chapter 5 The Relativistic Point Particle To formulate the dynamics of a system we can write either the equations of motion, or alternatively, an action. In the case of the relativistic point par- ticle, it is rather easy to write the equations of motion. But the action is so physical and geometrical that it is worth pursuing in its own right. More importantly, while it is difficult to guess the equations of motion for the rela- tivistic string, the action is a natural generalization of the relativistic particle action that we will study in this chapter. We conclude with a discussion of the charged relativistic particle. 5.1 Action for a relativistic point particle How can we find the action S that governs the dynamics of a free relativis- tic particle? To get started we first think about units. The action is the Lagrangian integrated over time, so the units of action are just the units of the Lagrangian multiplied by the units of time. The Lagrangian has units of energy, so the units of action are L2 ML2 [S]=M T = . (5.1.1) T 2 T Recall that the action Snr for a free non-relativistic particle is given by the time integral of the kinetic energy: 1 dx S = mv2(t) dt , v2 ≡ v · v, v = . (5.1.2) nr 2 dt 105 106 CHAPTER 5. THE RELATIVISTIC POINT PARTICLE The equation of motion following by Hamilton’s principle is dv =0. (5.1.3) dt The free particle moves with constant velocity and that is the end of the story. -

Chapter 5 ANGULAR MOMENTUM and ROTATIONS
Chapter 5 ANGULAR MOMENTUM AND ROTATIONS In classical mechanics the total angular momentum L~ of an isolated system about any …xed point is conserved. The existence of a conserved vector L~ associated with such a system is itself a consequence of the fact that the associated Hamiltonian (or Lagrangian) is invariant under rotations, i.e., if the coordinates and momenta of the entire system are rotated “rigidly” about some point, the energy of the system is unchanged and, more importantly, is the same function of the dynamical variables as it was before the rotation. Such a circumstance would not apply, e.g., to a system lying in an externally imposed gravitational …eld pointing in some speci…c direction. Thus, the invariance of an isolated system under rotations ultimately arises from the fact that, in the absence of external …elds of this sort, space is isotropic; it behaves the same way in all directions. Not surprisingly, therefore, in quantum mechanics the individual Cartesian com- ponents Li of the total angular momentum operator L~ of an isolated system are also constants of the motion. The di¤erent components of L~ are not, however, compatible quantum observables. Indeed, as we will see the operators representing the components of angular momentum along di¤erent directions do not generally commute with one an- other. Thus, the vector operator L~ is not, strictly speaking, an observable, since it does not have a complete basis of eigenstates (which would have to be simultaneous eigenstates of all of its non-commuting components). This lack of commutivity often seems, at …rst encounter, as somewhat of a nuisance but, in fact, it intimately re‡ects the underlying structure of the three dimensional space in which we are immersed, and has its source in the fact that rotations in three dimensions about di¤erent axes do not commute with one another. -

Chapter 3 Motion in Two and Three Dimensions
Chapter 3 Motion in Two and Three Dimensions 3.1 The Important Stuff 3.1.1 Position In three dimensions, the location of a particle is specified by its location vector, r: r = xi + yj + zk (3.1) If during a time interval ∆t the position vector of the particle changes from r1 to r2, the displacement ∆r for that time interval is ∆r = r1 − r2 (3.2) = (x2 − x1)i +(y2 − y1)j +(z2 − z1)k (3.3) 3.1.2 Velocity If a particle moves through a displacement ∆r in a time interval ∆t then its average velocity for that interval is ∆r ∆x ∆y ∆z v = = i + j + k (3.4) ∆t ∆t ∆t ∆t As before, a more interesting quantity is the instantaneous velocity v, which is the limit of the average velocity when we shrink the time interval ∆t to zero. It is the time derivative of the position vector r: dr v = (3.5) dt d = (xi + yj + zk) (3.6) dt dx dy dz = i + j + k (3.7) dt dt dt can be written: v = vxi + vyj + vzk (3.8) 51 52 CHAPTER 3. MOTION IN TWO AND THREE DIMENSIONS where dx dy dz v = v = v = (3.9) x dt y dt z dt The instantaneous velocity v of a particle is always tangent to the path of the particle. 3.1.3 Acceleration If a particle’s velocity changes by ∆v in a time period ∆t, the average acceleration a for that period is ∆v ∆v ∆v ∆v a = = x i + y j + z k (3.10) ∆t ∆t ∆t ∆t but a much more interesting quantity is the result of shrinking the period ∆t to zero, which gives us the instantaneous acceleration, a. -
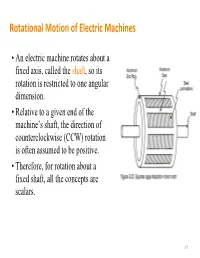
Rotational Motion of Electric Machines
Rotational Motion of Electric Machines • An electric machine rotates about a fixed axis, called the shaft, so its rotation is restricted to one angular dimension. • Relative to a given end of the machine’s shaft, the direction of counterclockwise (CCW) rotation is often assumed to be positive. • Therefore, for rotation about a fixed shaft, all the concepts are scalars. 17 Angular Position, Velocity and Acceleration • Angular position – The angle at which an object is oriented, measured from some arbitrary reference point – Unit: rad or deg – Analogy of the linear concept • Angular acceleration =d/dt of distance along a line. – The rate of change in angular • Angular velocity =d/dt velocity with respect to time – The rate of change in angular – Unit: rad/s2 position with respect to time • and >0 if the rotation is CCW – Unit: rad/s or r/min (revolutions • >0 if the absolute angular per minute or rpm for short) velocity is increasing in the CCW – Analogy of the concept of direction or decreasing in the velocity on a straight line. CW direction 18 Moment of Inertia (or Inertia) • Inertia depends on the mass and shape of the object (unit: kgm2) • A complex shape can be broken up into 2 or more of simple shapes Definition Two useful formulas mL2 m J J() RRRR22 12 3 1212 m 22 JRR()12 2 19 Torque and Change in Speed • Torque is equal to the product of the force and the perpendicular distance between the axis of rotation and the point of application of the force. T=Fr (Nm) T=0 T T=Fr • Newton’s Law of Rotation: Describes the relationship between the total torque applied to an object and its resulting angular acceleration. -

SPEED MANAGEMENT ACTION PLAN Implementation Steps
SPEED MANAGEMENT ACTION PLAN Implementation Steps Put Your Speed Management Plan into Action You did it! You recognized that speeding is a significant Figuring out how to implement a speed management plan safety problem in your jurisdiction, and you put a lot of time can be daunting, so the Federal Highway Administration and effort into developing a Speed Management Action (FHWA) has developed a set of steps that agency staff can Plan that holds great promise for reducing speeding-related adopt and tailor to get the ball rolling—but not speeding! crashes and fatalities. So…what’s next? Agencies can use these proven methods to jump start plan implementation and achieve success in reducing speed- related crashes. Involve Identify a Stakeholders Champion Prioritize Strategies for Set Goals, Track Implementation Market the Develop a Speed Progress, Plan Management Team Evaluate, and Celebrate Success Identify Strategy Leads INVOLVE STAKEHOLDERS In order for the plan to be successful, support and buy-in is • Outreach specialists. needed from every area of transportation: engineering (Federal, • Governor’s Highway Safety Office representatives. State, local, and Metropolitan Planning Organizations (MPO), • National Highway Transportation Safety Administration (NHTSA) Regional Office representatives. enforcement, education, and emergency medical services • Local/MPO/State Department of Transportation (DOT) (EMS). Notify and engage stakeholders who were instrumental representatives. in developing the plan and identify gaps in support. Potential • Police/enforcement representatives. stakeholders may include: • FHWA Division Safety Engineers. • Behavioral and infrastructure funding source representatives. • Agency Traffic Operations Engineers. • Judicial representatives. • Agency Safety Engineers. • EMS providers. • Agency Pedestrian/Bicycle Coordinators. • Educators. • Agency Pavement Design Engineers. -

Exploring Robotics Joel Kammet Supplemental Notes on Gear Ratios
CORC 3303 – Exploring Robotics Joel Kammet Supplemental notes on gear ratios, torque and speed Vocabulary SI (Système International d'Unités) – the metric system force torque axis moment arm acceleration gear ratio newton – Si unit of force meter – SI unit of distance newton-meter – SI unit of torque Torque Torque is a measure of the tendency of a force to rotate an object about some axis. A torque is meaningful only in relation to a particular axis, so we speak of the torque about the motor shaft, the torque about the axle, and so on. In order to produce torque, the force must act at some distance from the axis or pivot point. For example, a force applied at the end of a wrench handle to turn a nut around a screw located in the jaw at the other end of the wrench produces a torque about the screw. Similarly, a force applied at the circumference of a gear attached to an axle produces a torque about the axle. The perpendicular distance d from the line of force to the axis is called the moment arm. In the following diagram, the circle represents a gear of radius d. The dot in the center represents the axle (A). A force F is applied at the edge of the gear, tangentially. F d A Diagram 1 In this example, the radius of the gear is the moment arm. The force is acting along a tangent to the gear, so it is perpendicular to the radius. The amount of torque at A about the gear axle is defined as = F×d 1 We use the Greek letter Tau ( ) to represent torque. -

Moment of Inertia
MOMENT OF INERTIA The moment of inertia, also known as the mass moment of inertia, angular mass or rotational inertia, of a rigid body is a quantity that determines the torque needed for a desired angular acceleration about a rotational axis; similar to how mass determines the force needed for a desired acceleration. It depends on the body's mass distribution and the axis chosen, with larger moments requiring more torque to change the body's rotation rate. It is an extensive (additive) property: for a point mass the moment of inertia is simply the mass times the square of the perpendicular distance to the rotation axis. The moment of inertia of a rigid composite system is the sum of the moments of inertia of its component subsystems (all taken about the same axis). Its simplest definition is the second moment of mass with respect to distance from an axis. For bodies constrained to rotate in a plane, only their moment of inertia about an axis perpendicular to the plane, a scalar value, and matters. For bodies free to rotate in three dimensions, their moments can be described by a symmetric 3 × 3 matrix, with a set of mutually perpendicular principal axes for which this matrix is diagonal and torques around the axes act independently of each other. When a body is free to rotate around an axis, torque must be applied to change its angular momentum. The amount of torque needed to cause any given angular acceleration (the rate of change in angular velocity) is proportional to the moment of inertia of the body. -

Owner's Manual
OWNER’S MANUAL GET TO KNOW YOUR SYSTEM 1-877-DRY-TIME 3 7 9 8 4 6 3 basementdoctor.com TABLE OF CONTENTS IMPORTANT INFORMATION 1 YOUR SYSTEM 2 WARRANTIES 2 TROUBLESHOOTING 6 ANNUAL MAINTENANCE 8 WHAT TO EXPECT 9 PROFESSIONAL DEHUMIDIFIER 9 ® I-BEAM/FORCE 10 ® POWER BRACES 11 DRY BASEMENT TIPS 12 REFERRAL PROGRAM 15 IMPORTANT INFORMATION Please read the following information: Please allow new concrete to cure (dry) completely before returning your carpet or any other object to the repaired areas. 1 This normally takes 4-6 weeks, depending on conditions and time of year. Curing time may vary. You may experience some minor hairline cracking and dampness 2 with your new concrete. This is normal and does not affect the functionality of your new system. When installing carpet over the new concrete, nailing tack strips 3 is not recommended. This may cause your concrete to crack or shatter. Use Contractor Grade Liquid Nails. It is the responsibility of the Homeowner to keep sump pump discharge lines and downspouts (if applicable) free of roof 4 materials and leaves. If these lines should become clogged with external material, The Basement Doctor® can repair them at an additional charge. If we applied Basement Doctor® Coating to your walls: • This should not be painted over unless the paint contains an anti-microbial for it is the make-up of the coating that prohibits 5 mold growth. • This product may not cover all previous colors on your wall. • It is OK to panel or drywall over the Basement Doctor® Coating.