Yandi Aquatic Management Report
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Engineering & Mining Journal
Know-How | Performance | Reliability With MineView® and SmartFlow® Becker Mining Systems offers two comprehensive and scalable data management solutions for your Digital Mine. MineView® is a powerful state-of-the-art 3D SCADA system, that analyses incoming data from various mine equipment and visualises it in a 3D mine model. SmartFlow® takes Tagging & Tracking to a new level: collected asset data is centrally processed and smart software analytics allow for process optimization and improved safety. MINEVIEW BECKER MINING SYSTEMS AG We have been at the forefront of technology in Energy Distribution, Automation, Communication, Transportation and Roof Support since 1964. Together with our customers we create and deliver highest quality solutions and services to make operations run more profi tably, reliably and safely. For more information go to www.becker-mining.com/digitalmine Becker Mining is a trademark of Becker Mining Systems AG. © 2018 Becker Mining Systems AG or one of its affi liates. DECEMBER 2018 • VOL 219 • NUMBER 12 FEATURES China’s Miners Promote New Era of Openness and Cooperation Major reforms within the mining sector and the government will foster green mines at home and greater investment abroad ....................................42 Defeating the Deleterious Whether at the head of a circuit or scavenging tailings, today’s flotation innovations address challenges presented by declining grades, rising costs and aging plants ..................................................................................52 Staying on Top of -
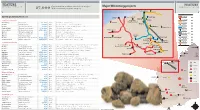
Feature Feature Major Wa Mining Projects
FEATURE FEATURE MINING OUTLOOK Construction workers needed for major Major WA mining projects MINING OUTLOOK 27,000 WA resources projects 2014-15 Source: Pitcrew Port Hedland Pardoo Rio Tinto rail MAJOR WA MINING PROJECTS Dampier Cape Lambert Iron Bridge Mt Dove Rio Tinto mine Completed in past year Balla Balla Abydos (Forge Resources) BHP rail Karara Mining Karara project $2.6bn Mid West Production ramp-up proceeding Sino Iron Wodgina Rio Tinto Hope Downs 4 mine $2.1bn Pilbara First production in H1 2013, ramping up to 15mtpa BHP mine Rio Tinto Marandoo mine expansion $1.1bn Pilbara Production will be sustained at 15mtpa for 16 further years Mt Webber McPhee Creek FMG rail Fortescue Metals Christmas Creek 2 expansion $US1.0bn Pilbara Completed in June 2013 quarter Fortescue Metals Port Hedland port expansion $US2.4bn Pilbara Fourth berth and support infrastructure opened in Aug 2013 Pannawonica FMG mine Atlas Iron Mt Dove mine development n/a Pilbara Production commenced in Dec 2012 Hancock proposed rail Atlas Iron Abydos mine development n/a Pilbara First haulage in Aug 2013, ramping up to 2-3mtpa Solomon Atlas Iron Utah Point 2 stockyard n/a Pilbara Largely complete and now ready to receive ore Hancock mine Hub Christmas Creek Rio Tinto Argyle Diamonds underground mine $US2.2bn Kimberley Production commenced in H1 2013 and is ramping up Buckland (Iron Ore Holdings) Cloudbreak Mineral Resources Sandfire Resources DeGrussa copper mine $US384m Mid West Ramp-up to nameplate production nearing completion Koodaideri Roy Hill Atlas Iron Construction -

Mining Tenement Summary Report
Government of Western Australia Department of Mines and Petroleum MINING TENEMENT SUMMARY REPORT MISCELLANEOUS LICENCE 52/163 Status: Live TENEMENT SUMMARY Area: 36.37000 HA Death Reason : Mark Out : N/A Death Date : Received : 02/09/2015 08:33:48 Commence : 09/03/2016 Term Granted : 21 Years Expiry : 08/03/2037 CURRENT HOLDER DETAILS Name and Address BHP BILLITON MINERALS PTY LTD WAIO LAND TENURE, C/- LAND TENURE TEAM, PO BOX 7474 CLOISTERS SQUARE, PERTH, WA, 6850, [email protected] ITOCHU MINERALS & ENERGY OF AUSTRALIA PTY LTD WAIO LAND TENURE, C/- LAND TENURE TEAM, PO BOX 7474, CLOISTERS SQUARE, PERTH, WA, 6850, [email protected] MITSUI-ITOCHU IRON PTY LTD WAIO LAND TENURE, C/- LAND TENURE TEAM, PO BOX 7474 CLOISTERS SQUARE, PERTH, WA, 6850, [email protected] DESCRIPTION Locality: Ophthalmia Dam Datum: Datum is at MGA94 Zone 50 Coordinates. 798661.887mE; 7415783.466mN Boundary: Thence 798661.887mE 7415783.466mN 798195.142mE 7414537.433mN 798145.514mE 7414505.938mN 797110.415mE 7414753.892mN 797115.012mE 7414725.174mN 796870.169mE 7414567.479mN 796849.420mE 7414561.946mN 796465.521mE 7414475.760mN 796462.363mE 7414475.051mN 796460.004mE 7414474.521mN 796399.464mE 7414460.930mN 796226.110mE 7414459.887mN 796050.820mE 7414470.256mN 796032.918mE 7414526.242mN 796068.190mE 7414559.056mN 796379.567mE 7414559.171mN 796736.615mE 7414657.382mN 796815.910mE 7414657.431mN 796886.689mE 7414688.863mN 797014.764mE 7414775.141mN 797075.629mE 7414824.500mN 797079.962mE 7414852.758mN 797112.933mE 7414856.133mN -

Notice of Meeting for Approval of the Proposed Transaction with BBIG
30 January 2020 ASX ANNOUNCEMENT Notice of Meeting for approval of the Proposed Transaction with BBIG Flinders Mines Limited (ASX:FMS) (Flinders) is pleased to announce the release of the attached notice of meeting, including an explanatory memorandum and independent expert's report (Notice of Meeting) in respect of an Extraordinary General Meeting (EGM) to consider the proposed transaction with BBI Group Pty Ltd (BBIG) to form an incorporated joint venture for the development of Flinders' Pilbara Iron Ore Project (PIOP), as announced on 28 November 2019 (Proposed Transaction). The Proposed Transaction represents the outcome of extensive commercial negotiations conducted by the Company’s independent PIOP Infrastructure Committee with BBIG to provide an infrastructure solution, facilitate the development of the PIOP and provide a pathway to market. Vote in favour of the Proposed Transaction The Independent Flinders Directors (Neil Warburton, The Hon. Cheryl Edwardes AM and James Gurry) unanimously recommend that Flinders shareholders vote in favour of the Proposed Transaction in the absence of a superior proposal. The Independent Flinders Directors appointed Grant Samuel as independent expert to consider and provide an opinion on the Proposed Transaction. The independent expert has concluded that the Proposed Transaction is fair and reasonable to non-associated Flinders shareholders, i.e. those shareholders other than TIO (NZ) Pty Ltd (TIO), Flinders’ largest shareholder and also a shareholder of BBIG. TIO will also be excluded from voting in favour of the Proposed Transaction. Ms Edwardes, Deputy Chair and Chair of Flinders’ PIOP Infrastructure Committee, said: “I am very pleased with the outcome of the negotiations with BBIG. -

Systematic Taxonomy and Biogeography of Widespread
Systematics and Biogeography of Three Widespread Australian Freshwater Fish Species. by Bernadette Mary Bostock B.Sc. (Hons) Submitted in fulfilment of the requirements for the degree of Doctor of Philosophy Deakin University February 2014 i ABSTRACT The variation within populations of three widespread and little studied Australian freshwater fish species was investigated using molecular genetic techniques. The three species that form the focus of this study are Leiopotherapon unicolor, Nematalosa erebi and Neosilurus hyrtlii, commonly recognised as the three most widespread Australian freshwater fish species, all are found in most of the major Australian drainage basins with habitats ranging from clear running water to near stagnant pools. This combination of a wide distribution and tolerance of a wide range of ecological conditions means that these species are ideally suited for use in investigating phylogenetic structure within and amongst Australian drainage basins. Furthermore, the combination of increasing aridity of the Australian continent and its diverse freshwater habitats is likely to promote population differentiation within freshwater species through the restriction of dispersal opportunities and localised adaptation. A combination of allozyme and mtDNA sequence data were employed to test the null hypothesis that Leiopotherapon unicolor represents a single widespread species. Conventional approaches to the delineation and identification of species and populations using allozyme data and a lineage-based approach using mitochondrial 16S rRNA sequences were employed. Apart from addressing the specific question of cryptic speciation versus high colonisation potential in widespread inland fishes, the unique status of L. unicolor as both Australia’s most widespread inland fish and most common desert fish also makes this a useful species to test the generality of current biogeographic hypotheses relating to the regionalisation of the Australian freshwater fish fauna. -

Other Land Users
25 YEARS OF NATIVE TITLE RECOGNITION Contents Overview 1 Argyle, Western Australia 2 Burrup Ancient Art 2 Rio Tinto agreements with 3 traditional owners Facilitating engagement on 3 cultural heritage Landmark High Court 3 decision: native title rights not extinguished by Mining Leases Native title railway 4 agreement to protect rock Bendigo Mining Agreement, signed in 2000 art Historic agreement: Pilbara 4 OTHER LAND USERS traditional owners and Rio Tinto Iron Ore The introduction of the Native Title Act 1993(Cth) had a profound effect on other land users, including the resource sector, pastoralists and graziers, Agreement to protect 5 fishers and local government. For the first time there was a legal requirement Gnulli heritage to take into account the native title rights and interests of Indigenous people The development of 6 when developing land. The initial response was one of fear and uncertainty; Australian Indigenous rights however, these sectors soon adapted to the new processes and became and Rio Tinto’s Indigenous regular participants in the native title process. involvement Watch: Ms Joanne Farrell, Managing Director Australia, Rio Tinto provide a perspective from the resource sector. OTHER LAND USERS Argyle, Western Australia Rio Tinto’s Argyle mine has a Participation Agreement in place with the Traditional Owners of the Argyle land to ensure that they benefit directly from the mine’s operations, now and for generations to come. This agreement —built on the principles of co-commitment, partnership and mutual trust—encompasses land rights, income generation, employment and contracting opportunities, land management and Indigenous site protection. A Traditional Owner relationship committee meets regularly to oversee the implementation of the agreement. -

Media Release
Media release INCREASE TO PILBARA ORE RESERVES AND MINERAL RESOURCES 6 March 2015 To support the annual Mineral Resources and Ore Reserves review process detailed in Rio Tinto’s 2014 Annual report released today, Rio Tinto Iron Ore has declared an increase of its managed Mineral Resources and Ore Reserves in the Pilbara, Western Australia, resulting from the completion of studies and evaluations. The update is reported under the Australasian Code for Reporting of Exploration Results, Mineral Resources and Ore Reserves, 2012 (JORC Code) and ASX Listing Rules, and provides a summary of information and Table 1 appendices to support the Mineral Resource Estimates and Ore Reserves for the material updates completed. Increases in Mineral Resources are reported as follows. 1. Hamersley Iron Brockman (increased by 681 million tonnes (Mt) to 2,998 Mt and Brockman process ore (increased by 324 Mt to 1,137 Mt) The increase represents minor changes across multiple deposits and the first inclusion of the Yandicoogina Braid deposit into the Mineral Resources, which was discovered by the Rio Tinto Exploration group during 2014. The Yandicoogina Braid deposit is classified as Inferred Mineral Resources and comprises: • 488 Mt at 61.7 per cent Fe Brockman Ore; • 238 Mt at 57.7 per cent Fe Brockman Process Ore; and • 62 Mt at 56.9 per cent Fe Channel Iron Deposit. 2. Robe Joint-Venture Channel Iron deposit This reflects an addition of 677 Mt of Inferred Mineral Resources at 53.1 per cent Fe of Channel Iron deposit mineralisation at the Jimmawurrada deposit, following metallurgical assessments of drill core. -

MAC – Yandi Rail Spur Inert Landfill Licence Application Supporting Documentation (Including Information Relating to Attachments 1 to 10)
MAC – Yandi Rail Spur Inert Landfill Licence Application Supporting Documentation (Including Information relating to Attachments 1 to 10) September 2018 Jimblebar L5415/1988/9 Licence Amendment Supporting Documentation (Including Information relating to Attachments 1 to 10) October 2017 MAC – Yandi Rail Spur Inert Landfill Licence Application Supporting Document Contents 1. Introduction ................................................................................................................................................................ 2 2. Project Description .................................................................................................................................................... 3 3. Existing Environment ................................................................................................................................................. 4 4. Environmental Management ...................................................................................................................................... 6 5. Environmental Impact Assessment and Associated Management Strategies ........................................................... 7 6. Heritage ................................................................................................................................................................... 11 7. Community Consultation ......................................................................................................................................... 11 8. Conclusion .............................................................................................................................................................. -

Arrium Limited Asx Appendix 4E Abn 63 004 410 833 Preliminary Final Report
ARRIUM LIMITED ASX APPENDIX 4E ABN 63 004 410 833 PRELIMINARY FINAL REPORT ARRIUM LIMITED RESULTS FOR ANNOUNCEMENT TO THE MARKET FINANCIAL YEAR ENDED 30 JUNE 2014 Comparison to previous corresponding period (pcp) Movement % Change A$ million Total revenue from ordinary activities up 2.4% to 7,006.6 Profit from ordinary activities after tax attributable up NC* to 205.4 to ordinary equity holders Net profit for the period attributable to ordinary up NC* to 205.4 equity holders * NC = Not comparable Dividends Final Dividend 2014 Interim Dividend 2014 Amount per security 3.0c 6.0c Franked amount per security 0.0c 0.0c Amount per security declared to be conduit 0.0c 0.0c foreign income Total dividend and dividend payment (A$ million) 41.0 81.7 Date Ex-dividend date for Final Dividend 10 September 2014 Record date for determining entitlement to Final Dividend 12 September 2014 Last day to elect to participate in the Dividend Reinvestment Plan (DRP) 15 September 2014 Date payable (on or around) 16 October 2014 Net tangible assets 30 June 2014 30 June 2013 Net Tangible Assets per security ($) 1.29 1.19 For personal use only ARRIUM LIMITED ASX APPENDIX 4E ABN 63 004 410 833 PRELIMINARY FINAL REPORT Details of Associates and Joint Venture Entities Name of associate or joint venture entity Associate or Joint Percentage Percentage Venture Entity holding holding June 2014 June 2013 BOSFA Pty Ltd Joint Venture 50% 50% GenAlta Recycling Inc. Joint Venture 50% 50% Details of entities over which control was gained during the period Name of entity Date Arrium Mining Sales Asia Limited 30 July 2013 Arrium Mining Services Asia Limited 26 July 2013 Moly-Cop Finance UK Limited 19 May 2014 There were no entities over which control was lost during the period. -

ASFIS ISSCAAP Fish List February 2007 Sorted on Scientific Name
ASFIS ISSCAAP Fish List Sorted on Scientific Name February 2007 Scientific name English Name French name Spanish Name Code Abalistes stellaris (Bloch & Schneider 1801) Starry triggerfish AJS Abbottina rivularis (Basilewsky 1855) Chinese false gudgeon ABB Ablabys binotatus (Peters 1855) Redskinfish ABW Ablennes hians (Valenciennes 1846) Flat needlefish Orphie plate Agujón sable BAF Aborichthys elongatus Hora 1921 ABE Abralia andamanika Goodrich 1898 BLK Abralia veranyi (Rüppell 1844) Verany's enope squid Encornet de Verany Enoploluria de Verany BLJ Abraliopsis pfefferi (Verany 1837) Pfeffer's enope squid Encornet de Pfeffer Enoploluria de Pfeffer BJF Abramis brama (Linnaeus 1758) Freshwater bream Brème d'eau douce Brema común FBM Abramis spp Freshwater breams nei Brèmes d'eau douce nca Bremas nep FBR Abramites eques (Steindachner 1878) ABQ Abudefduf luridus (Cuvier 1830) Canary damsel AUU Abudefduf saxatilis (Linnaeus 1758) Sergeant-major ABU Abyssobrotula galatheae Nielsen 1977 OAG Abyssocottus elochini Taliev 1955 AEZ Abythites lepidogenys (Smith & Radcliffe 1913) AHD Acanella spp Branched bamboo coral KQL Acanthacaris caeca (A. Milne Edwards 1881) Atlantic deep-sea lobster Langoustine arganelle Cigala de fondo NTK Acanthacaris tenuimana Bate 1888 Prickly deep-sea lobster Langoustine spinuleuse Cigala raspa NHI Acanthalburnus microlepis (De Filippi 1861) Blackbrow bleak AHL Acanthaphritis barbata (Okamura & Kishida 1963) NHT Acantharchus pomotis (Baird 1855) Mud sunfish AKP Acanthaxius caespitosa (Squires 1979) Deepwater mud lobster Langouste -

BHP OPERATIONAL REVIEW for the YEAR ENDED 30 JUNE 2021 Note: All Guidance Is Subject to Further Potential Impacts from COVID-19 During the 2022 Financial Year
Release Time IMMEDIATE Date 20 July 2021 Release Number 09/21 BHP OPERATIONAL REVIEW FOR THE YEAR ENDED 30 JUNE 2021 Note: All guidance is subject to further potential impacts from COVID-19 during the 2022 financial year. Record production was achieved at Western Australia Iron Ore (WAIO) and Goonyella. Olympic Dam achieved both the highest annual copper production since the acquisition by BHP in 2005 and the highest gold production ever for the operation. Escondida maintained average concentrator throughput at record levels despite a challenging operating environment in Chile as a result of impacts from COVID-19. Petroleum production for the 2021 financial year was slightly above guidance. Full year production guidance for copper, iron ore, metallurgical coal and nickel were delivered, as was revised guidance for energy coal. Full year unit cost guidance(1) expected to be achieved for WAIO, Escondida and Queensland Coal (based on exchange rates of AUD/USD 0.70 and USD/CLP 769). Petroleum unit costs are expected to be slightly better than guidance. New South Wales Energy Coal (NSWEC) unit costs are expected to be marginally above guidance. During the year, we successfully achieved first production at four major development projects, all of which were delivered on or ahead of schedule and on budget. The South Flank iron ore project in Western Australia and the Ruby oil and gas project in Trinidad and Tobago both achieved first production in May 2021. The Atlantis Phase 3 petroleum project and the Spence Growth Option copper project achieved first production in the first half of the 2021 financial year. -

Thatdeliver the Strategicdrivers
5041 BHPB AR06 cover_UK 13/9/06 10:35 PM Page 1 BHP Billiton Annual Report 2006 BHP Billiton Annual Report The Strategic Drivers that deliver the Essential Elements www.bhpbilliton.com Annual Report 2006 WorldReginfo - c6478d1e-7999-4617-a7c0-05343b86108a 5041 BHPB AR06 cover_UK 13/9/06 10:35 PM Page 2 Corporate Directory BHP BILLITON GROUP MARKETING OFFICES New Zealand We are BHP Billiton, a leading global resources REGISTERED OFFICES The Netherlands Computershare Investor Services Limited Level 2/159 Hurstmere Road company. BHP BILLITON LIMITED Verheeskade 25 2521 BE The Hague Takapuna North Shore City Australia Postal Address – Bag 92119 Auckland 1020 BHP Billiton Limited Telephone (31 70) 315 6666 Telephone (64 9) 488 8777 Our purpose is to create long-term value through the BHP Billiton Centre Facsimile (31 70) 315 6767 Facsimile (64 9) 488 8787 discovery, development and conversion of natural 180 Lonsdale Street Singapore Melbourne VIC 3000 168 Robinson Road #10-01 United States resources, and the provision of innovative customer Telephone (61 3) 9609 3333 Capital Tower Computershare Investor Services Facsimile (61 3) 9609 3015 Singapore 068912 2 North LaSalle Street and market-focused solutions. Telephone (65) 6349 3333 Chicago, IL 60602 BHP BILLITON PLC Facsimile (65) 6349 4000 Postal Address – PO Box 0289 United Kingdom Chicago, IL 60690-9569 Our seven strategic drivers assist us in achieving our Neathouse Place Telephone 1 888 404 6340 objectives. These drivers are our people; our licence to London SW1V 1BH SHARE REGISTRARS AND (toll-free within US) Telephone (44 20) 7802 4000 TRANSFER OFFICES Facsimile (1 312) 461 4331 operate; our world-class assets; the way we do business; Facsimile (44 20) 7802 4111 Australia ADR Depositary, Transfer Agent and Registrar our financial strength and discipline; our project pipeline; Company Secretaries BHP Billiton Limited Registrar JPMorgan Chase Bank, NA Computershare Investor Services JPMorgan Service Center Karen J Wood (Group Company Secretary) Pty Limited PO Box 3408 and growth options.