Canada Archives Canada Published Heritage Direction Du Branch Patrimoine De I'edition
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Namely, Aspartokinase and Aspartate 3-Semialdehyde Dehydrogenase) Are Not Subject to Feedback Inhibition Control by the End Product Methionine
REGULATION OF HOMOSERINE BIOSYNTHESIS BY L-CYSTEINE, A TERMINA L METABOLITE OF A LINKED PA THWA Y* By PRASANTA DATTA DEPARTMENT OF BIOLOGICAL CHEMISTRY, UNIVERSITY OF MICHIGAN, ANN ARBOR Communicated by Martin D. Kamen, June 23, 1967 In bacteria, the amino acid homoserine is a key branch-point intermediate in the synthesis of several amino acids of the aspartic pathway.' On the one hand, homoserine is converted to threonine (and thus to isoleucine), and through a separate sequence of reactions is transformed to methionine.1 In the latter se- quence, a succinylated product of homoserine is condensed with cysteine to pro- duce cystathionine,2-5 a precursor of methionine. Recent studies (see refs. 6 and 7 for up-to-date reviews on the subject) with several microorganisms have revealed that the synthesis of homoserine is regulated by various combinations of repression and/or feedback inhibition controls of early biosynthetic enzymes of the aspartic pathway by several end-product metabolites. One of these enzymes, homoserine dehydrogenase, catalyzes the pyridine nucleotide-linked reduction of aspartate j3-semialdehyde to homoserine. In the various bacteria that have been examined, this dehydrogenase as well as the two earlier enzymes of the path- way (namely, aspartokinase and aspartate 3-semialdehyde dehydrogenase) are not subject to feedback inhibition control by the end product methionine. This report describes the results of experiments on the control of activity of several bacterial homoserine dehydrogenases by L-cysteine, a terminal metabolite of a linked or connecting pathway, required for the synthesis of cystathionine.2-5 The findings suggest that the size of the homoserine pool in bacterial cells must depend, at least in part, on complex interdependent regulatory interactions of both cysteine and threonine on homoserine dehydrogenase. -

Joseph M. Jez • Department of Biology • Washington University in St
Joseph M. Jez • Department of Biology • Washington University in St. Louis Joseph M. Jez Professor and Howard Hughes Medical Institute Professor Department of Biology, Washington University in St. Louis One Brookings Drive, Campus Box 1137, St. Louis, MO 63130-4899 Phone: 314-935-3376; Email: [email protected] –––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––– EDUCATION University of Pennsylvania, Philadelphia, PA Ph.D. Biochemistry & Molecular Biophysics, 1998 Thesis: Steroid Recognition and Engineering of Catalysis in Mammalian Aldo-Keto Reductases Research Advisor: Prof. Trevor M. Penning Penn State University, University Park, PA B.S. Biochemistry (with Honors and English minor), 1992 Research Advisor: Prof. Gregory K. Farber PROFESSIONAL EXPERIENCE Washington University in St. Louis, St. Louis, MO Professor, Department of Biology (2015-current) Co-Director, Plant and Microbial Biosciences Program, Division of Biology & Biomedical Sciences (2013-current) Associate Professor, Department of Biology (2011-2015) Assistant Professor, Department of Biology (2008-2011) Honorary Assistant Professor, Department of Biology (2006-2008) Donald Danforth Plant Science Center, St. Louis, MO Assistant Member & Principal Investigator (2002-2010) Kosan Biosciences, Hayward, CA Scientist, New Technology Group (2001-2002) Supervisor: Dr. Daniel V. Santi The Salk Institute for Biological Studies, La Jolla, CA NIH-NRSA Postdoctoral Research Fellow, Structural Biology Laboratory (1998-2001) Project: Structure, Mechanism, -
When the Reaction Is
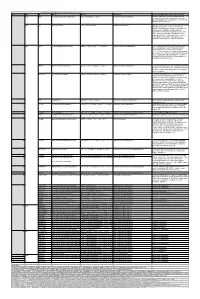
Table S3. iJL1678-ME model modification (blocked reactions) Iter. Cat. ID Name Formula Subsystem Comments (When the reaction is turned on) 1 bp2 EDD 6-phosphogluconate dehydratase 6pgc_c⇌2ddg6p_c + h2o_c Pentose Phosphate Pathway Create a major effect of steep acetate overflow elevation in high growth. Comparing to the main glycolytic pathway, it is metabolicly less efficient but proteomicly more efficient. bp1 ICL Isocitrate lyase icit_c→glx_c + succ_c Anaplerotic Reactions Bypass for the main TCA cycle pathways from turning isocitrate to succinate, when ICL is turned on, Isocitrate dehydrogenase(ICDHyr), 2-Oxogluterate dehydrogenase(AKGDH) and Succinyl-CoA synthetase (ATP-forming,SUCOAS) would reduce. Ref. (1) and (2) shows that this reaction is off in higher growth. Ref. (3) shows that this reaction is converging to being off when the dynamic of respiration using enzyme kinetics is simulated. 2 bp1 ABTA 4-aminobutyrate transaminase 4abut_c + akg_c⇌glu__L_c + sucsal_c Arginine and Proline Metabolism Another backup pathway of succinate production, from 2-Oxoglutarate (akg). Respiration would be induced when it is on, since the flux through ETC(CYTBO3_4pp and ATPS4rpp) would increase. As it requires the co-factor pyridoxal 5'-phosphate(2−) to get catalyzed(4), indicating that this reaction is regulated by the flux of other reactions(pyridoxal 5'- phosphate(2-) production, etc.). 3 GLYAT Glycine C-acetyltransferase accoa_c + gly_c⇌2aobut_c + coa_c Glycine and Serine Metabolism A reaction that back up for the respiration. Reactions fluxes in TCA cycle would drop when this reaction is turned on. It also requires pyridoxal 5'-phosphate(2−) for the regulation. 4 NADTRHD NAD transhydrogenase nad_c + nadph_c⇌nadh_c + nadp_c Oxidative Phosphorylation A reaction that would make the transition between NAD and NADP metabolically more efficient. -

Supplementary Table S4. FGA Co-Expressed Gene List in LUAD
Supplementary Table S4. FGA co-expressed gene list in LUAD tumors Symbol R Locus Description FGG 0.919 4q28 fibrinogen gamma chain FGL1 0.635 8p22 fibrinogen-like 1 SLC7A2 0.536 8p22 solute carrier family 7 (cationic amino acid transporter, y+ system), member 2 DUSP4 0.521 8p12-p11 dual specificity phosphatase 4 HAL 0.51 12q22-q24.1histidine ammonia-lyase PDE4D 0.499 5q12 phosphodiesterase 4D, cAMP-specific FURIN 0.497 15q26.1 furin (paired basic amino acid cleaving enzyme) CPS1 0.49 2q35 carbamoyl-phosphate synthase 1, mitochondrial TESC 0.478 12q24.22 tescalcin INHA 0.465 2q35 inhibin, alpha S100P 0.461 4p16 S100 calcium binding protein P VPS37A 0.447 8p22 vacuolar protein sorting 37 homolog A (S. cerevisiae) SLC16A14 0.447 2q36.3 solute carrier family 16, member 14 PPARGC1A 0.443 4p15.1 peroxisome proliferator-activated receptor gamma, coactivator 1 alpha SIK1 0.435 21q22.3 salt-inducible kinase 1 IRS2 0.434 13q34 insulin receptor substrate 2 RND1 0.433 12q12 Rho family GTPase 1 HGD 0.433 3q13.33 homogentisate 1,2-dioxygenase PTP4A1 0.432 6q12 protein tyrosine phosphatase type IVA, member 1 C8orf4 0.428 8p11.2 chromosome 8 open reading frame 4 DDC 0.427 7p12.2 dopa decarboxylase (aromatic L-amino acid decarboxylase) TACC2 0.427 10q26 transforming, acidic coiled-coil containing protein 2 MUC13 0.422 3q21.2 mucin 13, cell surface associated C5 0.412 9q33-q34 complement component 5 NR4A2 0.412 2q22-q23 nuclear receptor subfamily 4, group A, member 2 EYS 0.411 6q12 eyes shut homolog (Drosophila) GPX2 0.406 14q24.1 glutathione peroxidase -

Supplementary Table 1
Supplementary Table 1. 492 genes are unique to 0 h post-heat timepoint. The name, p-value, fold change, location and family of each gene are indicated. Genes were filtered for an absolute value log2 ration 1.5 and a significance value of p ≤ 0.05. Symbol p-value Log Gene Name Location Family Ratio ABCA13 1.87E-02 3.292 ATP-binding cassette, sub-family unknown transporter A (ABC1), member 13 ABCB1 1.93E-02 −1.819 ATP-binding cassette, sub-family Plasma transporter B (MDR/TAP), member 1 Membrane ABCC3 2.83E-02 2.016 ATP-binding cassette, sub-family Plasma transporter C (CFTR/MRP), member 3 Membrane ABHD6 7.79E-03 −2.717 abhydrolase domain containing 6 Cytoplasm enzyme ACAT1 4.10E-02 3.009 acetyl-CoA acetyltransferase 1 Cytoplasm enzyme ACBD4 2.66E-03 1.722 acyl-CoA binding domain unknown other containing 4 ACSL5 1.86E-02 −2.876 acyl-CoA synthetase long-chain Cytoplasm enzyme family member 5 ADAM23 3.33E-02 −3.008 ADAM metallopeptidase domain Plasma peptidase 23 Membrane ADAM29 5.58E-03 3.463 ADAM metallopeptidase domain Plasma peptidase 29 Membrane ADAMTS17 2.67E-04 3.051 ADAM metallopeptidase with Extracellular other thrombospondin type 1 motif, 17 Space ADCYAP1R1 1.20E-02 1.848 adenylate cyclase activating Plasma G-protein polypeptide 1 (pituitary) receptor Membrane coupled type I receptor ADH6 (includes 4.02E-02 −1.845 alcohol dehydrogenase 6 (class Cytoplasm enzyme EG:130) V) AHSA2 1.54E-04 −1.6 AHA1, activator of heat shock unknown other 90kDa protein ATPase homolog 2 (yeast) AK5 3.32E-02 1.658 adenylate kinase 5 Cytoplasm kinase AK7 -

Construction of a Synthetic Metabolic Pathway for the Production of 2,4
Construction of a synthetic metabolic pathway for the production of 2,4-dihydroxybutyric acid from homoserine Thomas Walther, Florence Calvayrac, Yoann Malbert, Ceren Alkim, Clémentine Dressaire, Hélène Cordier, Jean Marie François To cite this version: Thomas Walther, Florence Calvayrac, Yoann Malbert, Ceren Alkim, Clémentine Dressaire, et al.. Construction of a synthetic metabolic pathway for the production of 2,4-dihydroxybutyric acid from homoserine. Metabolic Engineering, Elsevier, 2018, 10.1016/j.ymben.2017.12.005. hal-01886438 HAL Id: hal-01886438 https://hal.archives-ouvertes.fr/hal-01886438 Submitted on 27 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributed under a Creative Commons Attribution| 4.0 International License Author’s Accepted Manuscript Construction of a synthetic metabolic pathway for the production of 2,4-dihydroxybutyric acid from homoserine Thomas Walther, Florence Calvayrac, Yoann Malbert, Ceren Alkim, Clémentine Dressaire, Hélène Cordier, Jean Marie François www.elsevier.com/locate/ymben PII: S1096-7176(17)30350-6 -

Severe Child Form of Primary Hyperoxaluria Type 2
Konkoľová et al. BMC Medical Genetics (2017) 18:59 DOI 10.1186/s12881-017-0421-8 CASE REPORT Open Access Severe child form of primary hyperoxaluria type 2 - a case report revealing consequence of GRHPR deficiency on metabolism Jana Konkoľová1,2*, Ján Chandoga1,2, Juraj Kováčik3, Marcel Repiský2, Veronika Kramarová2, Ivana Paučinová3 and Daniel Böhmer1,2 Abstract Background: Primary hyperoxaluria type 2 is a rare monogenic disorder inherited in an autosomal recessive pattern. It results from the absence of the enzyme glyoxylate reductase/hydroxypyruvate reductase (GRHPR). As a consequence of deficient enzyme activity, excessive amounts of oxalate and L-glycerate are excreted in the urine, and are a source for the formation of calcium oxalate stones that result in recurrent nephrolithiasis and less frequently nephrocalcinosis. Case presentation: We report a case of a 10-month-old patient diagnosed with urolithiasis. Screening of inborn errors of metabolism, including the performance of GC/MS urine organic acid profiling and HPLC amino acid profiling, showed abnormalities, which suggested deficiency of GRHPR enzyme. Additional metabolic disturbances observed in the patient led us to seek other genetic determinants and the elucidation of these findings. Besides the elevated excretion of 3-OH-butyrate, adipic acid, which are typical marks of ketosis, other metabolites such as 3- aminoisobutyric acid, 3-hydroxyisobutyric acid, 3-hydroxypropionic acid and 2-ethyl-3-hydroxypropionic acids were observed in increased amounts in the urine. Direct sequencing of the GRHPR gene revealed novel mutation, described for the first time in this article c.454dup (p.Thr152Asnfs*39) in homozygous form. The frequent nucleotide variants were found in AGXT2 gene. -

Genomic Evidence of Reactive Oxygen Species Elevation in Papillary Thyroid Carcinoma with Hashimoto Thyroiditis
Endocrine Journal 2015, 62 (10), 857-877 Original Genomic evidence of reactive oxygen species elevation in papillary thyroid carcinoma with Hashimoto thyroiditis Jin Wook Yi1), 2), Ji Yeon Park1), Ji-Youn Sung1), 3), Sang Hyuk Kwak1), 4), Jihan Yu1), 5), Ji Hyun Chang1), 6), Jo-Heon Kim1), 7), Sang Yun Ha1), 8), Eun Kyung Paik1), 9), Woo Seung Lee1), Su-Jin Kim2), Kyu Eun Lee2)* and Ju Han Kim1)* 1) Division of Biomedical Informatics, Seoul National University College of Medicine, Seoul, Korea 2) Department of Surgery, Seoul National University Hospital and College of Medicine, Seoul, Korea 3) Department of Pathology, Kyung Hee University Hospital, Kyung Hee University School of Medicine, Seoul, Korea 4) Kwak Clinic, Okcheon-gun, Chungbuk, Korea 5) Department of Internal Medicine, Uijeongbu St. Mary’s Hospital, Uijeongbu, Korea 6) Department of Radiation Oncology, Seoul St. Mary’s Hospital, Seoul, Korea 7) Department of Pathology, Chonnam National University Hospital, Kwang-Ju, Korea 8) Department of Pathology, Samsung Medical Center, Sungkyunkwan University School of Medicine, Seoul, Korea 9) Department of Radiation Oncology, Korea Cancer Center Hospital, Korea Institute of Radiological and Medical Sciences, Seoul, Korea Abstract. Elevated levels of reactive oxygen species (ROS) have been proposed as a risk factor for the development of papillary thyroid carcinoma (PTC) in patients with Hashimoto thyroiditis (HT). However, it has yet to be proven that the total levels of ROS are sufficiently increased to contribute to carcinogenesis. We hypothesized that if the ROS levels were increased in HT, ROS-related genes would also be differently expressed in PTC with HT. To find differentially expressed genes (DEGs) we analyzed data from the Cancer Genomic Atlas, gene expression data from RNA sequencing: 33 from normal thyroid tissue, 232 from PTC without HT, and 60 from PTC with HT. -

Identification of Six Novel Allosteric Effectors of Arabidopsis Thaliana Aspartate Kinase-Homoserine Dehydrogenase
Identification of six novel allosteric effectors of Arabidopsis thaliana Aspartate kinase-Homoserine dehydrogenase. Physiological context sets the specificity Gilles Curien, Stéphane Ravanel, Mylène Robert, Renaud Dumas To cite this version: Gilles Curien, Stéphane Ravanel, Mylène Robert, Renaud Dumas. Identification of six novel allosteric effectors of Arabidopsis thaliana Aspartate kinase-Homoserine dehydrogenase. Physiological context sets the specificity. Journal of Biological Chemistry, American Society for Biochemistry and Molecular Biology, 2005, 280 (50), pp.41178-41183. 10.1074/jbc.M509324200. hal-00016689 HAL Id: hal-00016689 https://hal.archives-ouvertes.fr/hal-00016689 Submitted on 31 May 2020 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 280, NO. 50, pp. 41178–41183, December 16, 2005 © 2005 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. Identification of Six Novel Allosteric Effectors of Arabidopsis thaliana Aspartate Kinase-Homoserine Dehydrogenase -

Molecular Changes in Tissue Proteome During Prostate Cancer Development: Proof-Of-Principle Investigation
diagnostics Article Molecular Changes in Tissue Proteome during Prostate Cancer Development: Proof-of-Principle Investigation Agnieszka Latosinska 1,*, Katarina Davalieva 2 , Manousos Makridakis 3, William Mullen 4 , Joost P. Schanstra 5,6, Antonia Vlahou 3, Harald Mischak 1 and Maria Frantzi 1 1 Mosaiques Diagnostics GmbH, 30659 Hannover, Germany; [email protected] (H.M.); [email protected] (M.F.) 2 Research Centre for Genetic Engineering and Biotechnology “Georgi D Efremov”, Macedonian Academy of Sciences and Arts, 1000 Skopje, North Macedonia; [email protected] 3 Biomedical Research Foundation of the Academy of Athens, Centre of Systems Biology, 11527 Athens, Greece; [email protected] (M.M.); [email protected] (A.V.) 4 Institute of Cardiovascular and Medical Sciences, University of Glasgow, Glasgow G12 8QQ, UK; [email protected] 5 Institut National de la Santé et de la Recherche Médicale (INSERM), U1048, Institute of Cardiovascular and Metabolic Diseases, 31432 Toulouse, France; [email protected] 6 Université Toulouse III Paul-Sabatier, 31400 Toulouse, France * Correspondence: [email protected]; Tel.: +49-(0)51-15-547-4430; Fax: +49-(0)51-15-547-4431 Received: 30 July 2020; Accepted: 26 August 2020; Published: 31 August 2020 Abstract: (1) Background: Prostate cancer (PCa) is characterized by high heterogeneity. The aim of this study was to investigate molecular alterations underlying PCa development based on proteomics data. (2) Methods: Liquid chromatography coupled to tandem mass spectrometry was conducted for 22 fresh-frozen tissue specimens from patients with benign prostatic hyperplasia (BPH, n = 5) and PCa (n = 17). Mann Whitney test was used to define significant differences between the two groups. -

Phosphoglycolate Salvage in a Chemolithoautotroph Using the Calvin Cycle
Phosphoglycolate salvage in a chemolithoautotroph using the Calvin cycle Nico J. Claassensa,1, Giovanni Scarincia,1, Axel Fischera, Avi I. Flamholzb, William Newella, Stefan Frielingsdorfc, Oliver Lenzc, and Arren Bar-Evena,2 aSystems and Synthetic Metabolism Lab, Max Planck Institute of Molecular Plant Physiology, 14476 Potsdam-Golm, Germany; bDepartment of Molecular and Cell Biology, University of California, Berkeley, CA 94720; and cInstitut für Chemie, Physikalische Chemie, Technische Universität Berlin, 10623 Berlin, Germany Edited by Donald R. Ort, University of Illinois at Urbana–Champaign, Urbana, IL, and approved July 24, 2020 (received for review June 14, 2020) Carbon fixation via the Calvin cycle is constrained by the side In the cyanobacterium Synechocystis sp. PCC6803, gene dele- activity of Rubisco with dioxygen, generating 2-phosphoglycolate. tion studies were used to demonstrate the activity of two pho- The metabolic recycling of phosphoglycolate was extensively torespiratory routes in addition to the C2 cycle (5, 8). In the studied in photoautotrophic organisms, including plants, algae, glycerate pathway, two glyoxylate molecules are condensed to and cyanobacteria, where it is referred to as photorespiration. tartronate semialdehyde, which is subsequently reduced to glyc- While receiving little attention so far, aerobic chemolithoautotro- erate and phosphorylated to 3PG (Fig. 1). Alternatively, in the phic bacteria that operate the Calvin cycle independent of light oxalate decarboxylation pathway, glyoxylate is oxidized -

Genome-Scale Metabolic Network Analysis and Drug Targeting of Multi-Drug Resistant Pathogen Acinetobacter Baumannii AYE
Electronic Supplementary Material (ESI) for Molecular BioSystems. This journal is © The Royal Society of Chemistry 2017 Electronic Supplementary Information (ESI) for Molecular BioSystems Genome-scale metabolic network analysis and drug targeting of multi-drug resistant pathogen Acinetobacter baumannii AYE Hyun Uk Kim, Tae Yong Kim and Sang Yup Lee* E-mail: [email protected] Supplementary Table 1. Metabolic reactions of AbyMBEL891 with information on their genes and enzymes. Supplementary Table 2. Metabolites participating in reactions of AbyMBEL891. Supplementary Table 3. Biomass composition of Acinetobacter baumannii. Supplementary Table 4. List of 246 essential reactions predicted under minimal medium with succinate as a sole carbon source. Supplementary Table 5. List of 681 reactions considered for comparison of their essentiality in AbyMBEL891 with those from Acinetobacter baylyi ADP1. Supplementary Table 6. List of 162 essential reactions predicted under arbitrary complex medium. Supplementary Table 7. List of 211 essential metabolites predicted under arbitrary complex medium. AbyMBEL891.sbml Genome-scale metabolic model of Acinetobacter baumannii AYE, AbyMBEL891, is available as a separate file in the format of Systems Biology Markup Language (SBML) version 2. Supplementary Table 1. Metabolic reactions of AbyMBEL891 with information on their genes and enzymes. Highlighed (yellow) reactions indicate that they are not assigned with genes. No. Metabolism EC Number ORF Reaction Enzyme R001 Glycolysis/ Gluconeogenesis 5.1.3.3 ABAYE2829