Spatiotemporal Variation in Abundance and Social Structure of Bottlenose Dolphins in the Florida Coastal Everglades Robin E
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2019 Preliminary Manatee Mortality Table with 5-Year Summary From: 01/01/2019 To: 11/22/2019
FLORIDA FISH AND WILDLIFE CONSERVATION COMMISSION MARINE MAMMAL PATHOBIOLOGY LABORATORY 2019 Preliminary Manatee Mortality Table with 5-Year Summary From: 01/01/2019 To: 11/22/2019 County Date Field ID Sex Size Waterway City Probable Cause (cm) Nassau 01/01/2019 MNE19001 M 275 Nassau River Yulee Natural: Cold Stress Hillsborough 01/01/2019 MNW19001 M 221 Hillsborough Bay Apollo Beach Natural: Cold Stress Monroe 01/01/2019 MSW19001 M 275 Florida Bay Flamingo Undetermined: Other Lee 01/01/2019 MSW19002 M 170 Caloosahatchee River North Fort Myers Verified: Not Recovered Manatee 01/02/2019 MNW19002 M 213 Braden River Bradenton Natural: Cold Stress Putnam 01/03/2019 MNE19002 M 175 Lake Ocklawaha Palatka Undetermined: Too Decomposed Broward 01/03/2019 MSE19001 M 246 North Fork New River Fort Lauderdale Natural: Cold Stress Volusia 01/04/2019 MEC19002 U 275 Mosquito Lagoon Oak Hill Undetermined: Too Decomposed St. Lucie 01/04/2019 MSE19002 F 226 Indian River Fort Pierce Natural: Cold Stress Lee 01/04/2019 MSW19003 F 264 Whiskey Creek Fort Myers Human Related: Watercraft Collision Lee 01/04/2019 MSW19004 F 285 Mullock Creek Fort Myers Undetermined: Too Decomposed Citrus 01/07/2019 MNW19003 M 275 Gulf of Mexico Crystal River Verified: Not Recovered Collier 01/07/2019 MSW19005 M 270 Factory Bay Marco Island Natural: Other Lee 01/07/2019 MSW19006 U 245 Pine Island Sound Bokeelia Verified: Not Recovered Lee 01/08/2019 MSW19007 M 254 Matlacha Pass Matlacha Human Related: Watercraft Collision Citrus 01/09/2019 MNW19004 F 245 Homosassa River Homosassa -

Interrelationships Among Hydrological, Biodiversity and Land Use Features of the Pantanal and Everglades
Interrelationships among hydrological, biodiversity and Land Use Features of the Pantanal and Everglades Biogeochemical Segmentation and Derivation of Numeric Nutrient Criteria for Coastal Everglades waters. FIU Henry Briceño. Joseph N. Boyer NPS Joffre Castro 100 years of hydrology intervention …urban development 1953 1999 Naples Bay impacted by drainage, channelization, and urban development FDEP 2010 SEGMENTATION METHOD Six basins, 350 stations POR 1991 (1995)-1998. NH4, NO2, TOC, TP, TN, NO3, TON, SRP, DO, Turbidity, Salinity, CHLa, Temperature Factor Analysis (PC extraction) Scores Mean, SD, Median, MAD Hierarchical Clustering NUMERIC NUTRIENT CRITERIA The USEPA recommends three types of approaches for setting numeric nutrient criteria: - reference condition approach - stressor-response analysis - mechanistic modeling. A Station’s Never to Exceed (NTE) Limit. This limit is the highest possible level that a station concentration can reach at any time A Segment’s Annual Geometric Mean (AGM) Limit. This limit is the highest possible level a segment’s average concentration of annual geometric means can reach in year A Segment’s 1-in-3 Years (1in3) Limit. This limit is the level that a segment average concentration of annual geometric means should be less than or equal to, at least, twice in three consecutive years. 1in3 AGM NTE 90% 80% 95% AGM : Annual Geometric Mean Not to be exceeded 1in3 : Annual Geometric Mean Not to exceed more than once in 3 yrs Biscayne Bay, Annual Geometric Means 0.7 AGMAGM Limit : Not to be exceeded 0.6 (Annual Geometric Mean not to be exceeded) 1in31in3 Limit : Not to exceed more 0.5 (Annualthan Geometric once Mean in not 3 to beyears exceeded more than once in 3 yrs) 0.4 0.3 Total Nitrogen, mg/LNitrogen, Total 0.2 Potentially Enriched 0.1 SCO NCO SNB NCI NNB CS SCM SCI MBS THRESHOLD ANALYSIS Regime Shift Detection methods (Rodionov 2004) Cumulative deviations from mean method CTZ CHLa Zcusum Threshold 20 0 -20 Cusum . -

Watercraft Access Facilities in the State of Florida Biological Assessment
Watercraft Access Facilities in the State of Florida Biological Assessment Introduction The U.S. Army Corps of Engineers (Corps) issues numerous permits for watercraft access facilities in Florida manatee (Trichechus manatus latirostris) habitat within Florida. Permitting actions that involve the rehabilitation of existing facilities and the construction of new facilities that include 4 slips or less in manatee habitat are currently evaluated using a programmatic approach that meets the consultation requirements of the Endangered Species Act. The purpose of this action is to develop and implement a programmatic framework to encompass the majority of proposals (with the exception of proposals that involve the rehabilitation of existing facilities and the construction of new facilities that include 4 slips or less in manatee habitat), regardless of size or extent, in order to streamline the formal consultation process and ensure that adequate measures are in place to avoid and minimize impacts to manatees, as well as any adverse modification(s) of critical habitat. Project Description / Area of Analysis The actions addressed by this Biological Opinion include all new watercraft access facilities (i.e., docks, boat ramps, and marinas and any dredging, pipes and culverts included in project plans and build out) which are likely to adversely affect manatees as determined via the 2011 Manatee Key. This analysis is intended to assist biologists in assessing the potential impacts of watercraft access facilities on the Florida manatee (Trichechus manatus latirostris) in the State of Florida. Manatee Occurrence Florida manatees are found throughout the southeastern United States. As a subspecies of the West Indian manatee, their presence here represents the northern limit of this species range (Lefebvre et al. -
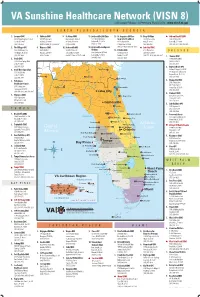
Veterans-Ride-Program-VA-Map.Pdf
Chatuge L. 59 Stevenson 76 Florence Athens 11 19 L. Burton Muscle Shoals 76 27 75 Scottsboro 123 129 Decatur Moulton 23 Hartselle Fort Payne 85 41 575 441 29 Albertville 19 129 Cullman Allatoona L. Hamilton Gadsden Kennesaw Mountain NBP Guin 278 Oneonta 29 27 Chattahoochee River NRA Sulligent Jasper 378 78 Sumiton Saks 20 Center Point Anniston 278 78 20 Mountain Brook 27A 29 129 278 Hueytown Talladega Reform VA Sunshine HealthcareSouth Augusta Network (VISN 8) Jackson L. 221 Alabaster 140 Fountain Parkway • St. Petersburg, Florida 33716 • www.visn8.va.gov Sylacauga 41 Calera Roanoke 75 1 23 27A BIBB Alexander City Brent West Point L. 25 COOSA UPSON WASHINGTON SCREVEN 441 Lafayette NORTHMONROE FLORIDA/SOUTH GEORGIA Eutaw TALLAPOOSA Macon Clanton CHAMBERS HARRIS BIBB Flint R. 80 JENKINS 19 319 TALBOT CHILTON 27 PERRY WILKINSON CRAWFORD JOHNSON 301 1. Lecanto CBOC 7. AuburnValdosta CBOC 9. St. Marys CBOC 11.LAUREN JacksonvilleS 2 VA Clinic 13. St. Augustine VA Clinic 15. Perry VA Clinic Malcom Randall VAMC 341 TAYLOR L. Harding 23 Oconee R. PEACH LEE TWIGGS 80 Demopolis 2804 W Marc Knighton Ct, SteELMORE A 2841 N Patterson St MUSCOGEE 2603 Osbourne Rd Ste E 3901 UniversityEMANUEL Blvd S (new interim address) 1224 N Peacock Ave 1601 SW Archer Rd 129 AUTAUGA Prattville HOUSTON BLECKLEY Selma Phenix City 16 Lecanto, FL 34461 Valdosta, GA 31601 Columbus St. Marys, GA 31558 Jacksonville, FL 32216 CANDLER 195EFFINGHAM Southpark Blvd Perry, FL 32347 Gainesville, FL 32608 MACON TREUTLEN CHATAHOOCHEE MACON BULLOCH 80 Linden (352)Montgomery 746-8000 (229) 293-0132;RUSSELL (877) 303-8387MARION (912) 510-3420 PULASKI (904) 732-6300 St Augustine, FL 32086 (850) 223-8387 (352) 376-1611; (800) 324-8387 221 SCHLEY 341 25 BULLOCK Andersonville NHS 41 95 LOWNDES Union Springs MONT- (904) 829-0814; (866) 401-8387 2. -

Bookletchart™ Florida Everglades National Park – Whitewater Bay NOAA Chart 11433
BookletChart™ Florida Everglades National Park – Whitewater Bay NOAA Chart 11433 A reduced-scale NOAA nautical chart for small boaters When possible, use the full-size NOAA chart for navigation. Included Area Published by the to Coot Bay and Whitewater Bay. A highway bridge, about 0.5 mile above the mouth of the canal, has a reported 45-foot fixed span and a National Oceanic and Atmospheric Administration clearance of 10 feet. A marina on the W side of the canal just below the National Ocean Service dam at Flamingo has berths with electricity, water, ice, and limited Office of Coast Survey marine supplies. Gasoline, diesel fuel, and launching ramps are available on either side of the dam. A 5-mph no-wake speed limit is enforced in www.NauticalCharts.NOAA.gov the canal. 888-990-NOAA Small craft can traverse the system of tidal bays, creeks, and canals from Flamingo Visitors Center to the Gulf of Mexico, 6 miles N of Northwest What are Nautical Charts? Cape. The route through Buttonwood Canal, Coot Bay, Tarpon Creek, Whitewater Bay, Cormorant Pass, Oyster Bay, and Little Shark River is Nautical charts are a fundamental tool of marine navigation. They show marked by daybeacons. The controlling depth is about 3½ feet. water depths, obstructions, buoys, other aids to navigation, and much The route from Flamingo to Daybeacon 48, near the W end of more. The information is shown in a way that promotes safe and Cormorant Pass, is part of the Wilderness Waterway. efficient navigation. Chart carriage is mandatory on the commercial Wilderness Waterway is a 100-mile inside passage winding through the ships that carry America’s commerce. -

Of 6 62-302.532 Estuary-Specific Numeric Interpretations of The
FAC 62-302.532 Estuary-Specific Numeric Interpretations of the Narrative Nutrient Criterion Effective Date: 12/20/2012 62-302.532 Estuary-Specific Numeric Interpretations of the Narrative Nutrient Criterion. (1) Estuary-specific numeric interpretations of the narrative nutrient criterion in paragraph 62-302.530(47)(b), F.A.C., are in the table below. The concentration-based estuary interpretations are open water, area-wide averages. The interpretations expressed as load per million cubic meters of freshwater inflow are the total load of that nutrient to the estuary divided by the total volume of freshwater inflow to that estuary. Page 1 of 6 FAC 62-302.532 Estuary-Specific Numeric Interpretations of the Narrative Nutrient Criterion Effective Date: 12/20/2012 Estuary Total Phosphorus Total Nitrogen Chlorophyll a (a) Clearwater Harbor/St. Joseph Sound Annual geometric mean values not to be exceeded more than once in a three year period. Nutrient and nutrient response values do not apply to tidally influenced areas that fluctuate between predominantly marine and predominantly fresh waters during typical climatic and hydrologic conditions. 1. St.Joseph Sound 0.05 mg/L 0.66 mg/L 3.1 µg/L 2. Clearwater North 0.05 mg/L 0.61 mg/L 5.4 µg/L 3. Clearwater South 0.06 mg/L 0.58 mg/L 7.6 µg/L (b) Tampa Bay Annual totals for nutrients and annual arithmetic means for chlorophyll a, not to be exceeded more than once in a three year period. Nutrient and nutrient response values do not apply to tidally influenced areas that fluctuate between predominantly marine and predominantly fresh waters during typical climatic and hydrologic conditions. -

25 June 2005 Kim Hanes SFWMD 8894 Belvedere Road West Palm Beach, FL 33411
Southeast Environmental Research Center OE-148 Florida International University, Miami, FL 33199 305-348-3095, 305-348-4096 fax, http://serc.fiu.edu 25 June 2005 Kim Hanes SFWMD 8894 Belvedere Road West Palm Beach, FL 33411 Re: South Florida Coastal Water Quality Monitoring Network – 1-3/05 Quarterly Report (C-15397) Dear Mr. Hanes: This letter serves to transmit the South Florida Coastal Water Quality Monitoring Network Quarterly Report as per our SFWMD/SERC Cooperative Agreement #C-15397. This report consists of this letter along with corresponding tables and figures. Project Background This report includes water quality data collected monthly during the annual period of record (POR) Jan. – Mar. 2005 from 28 stations in Florida Bay, 22 stations in Whitewater Bay, 25 stations in Ten Thousand Islands, 25 stations in Biscayne Bay, and 28 stations in Cape Romano-Rookery Bay-Pine Island Sound. A total of 49 stations were also collected on the SW Florida Shelf on a quarterly basis. Figure 1 shows the location of the fixed sampling stations. Water quality parameters monitored at each station include the dissolved nutrients nitrate + nitrite (NOx), nitrite (NO2), nitrate (NO3), ammonium (NH4), inorganic nitrogen (DIN), and soluble reactive phosphorus (SRP). Silicate (Si(OH)4) was analyzed at all stations on a quarterly basis in conjunction with SW Shelf sampling. Total concentrations of nitrogen (TN), organic nitrogen (TON), phosphorus (TP), and organic carbon (TOC) were also measured. All concentrations for each of these parameters are reported as parts per million (ppm) except where noted. Biological parameters monitored included chlorophyll a (µg l-1) and alkaline phosphatase activity (APA; µM hr-1). -

DISTRIBUTION and SALINITY TOLERANCE in the AMPHIURID BRITTLESTAR, OPHIOPHRAGMUS FILOGRANEUS (LYMAN, 187Sr
DISTRIBUTION AND SALINITY TOLERANCE IN THE AMPHIURID BRITTLESTAR, OPHIOPHRAGMUS FILOGRANEUS (LYMAN, 187Sr LOWELL P. THOMAS The Marine Laboratory, University of Miami ABSTRACT A brief discussion of the ecology and distribution of Ophiophragmus filograneus (Lyman) is given. This species is reported from salinities as low as 7.7%0' apparently a record low for echinoderms. Other examples of estuarine echinoderms are discussed. The amphiurid brittlestar Ophiophragmus filograneus (Lyman, 1875) was originally described from the Cedar Keys region of the west coast of Florida. Since its description this species has been re- ported only once, when Pearson (1936) found it in Biscayne Bay near Miami. The writer has collected O. filograneus on the Florida east coast as far north as Melbourne where it occurs in the Indian River, and as far south as Whitewater Bay in the Everglades National Park. Other specimens were taken at Ft. Myers and Marco on the west coast and Lake Worth on the east coast. In all cases they were captur- ed in the soft mud of bays separated from the ocean by land masses. Generally O. filograneus is associated with the marine monocotyledon Diplanthera wrightii (Ascherson) which has recently been treated by Phillips (1960). An apparently closely related ophiuroid, Ophio- phragmus wurdemani Lyman occurs in fine quartz sand of shallow, more saline waters along Gulf of Mexico and Atlantic coast beaches. While taking bottom samples from eastern Whitewater Bay during the summer of 1958 in connection with an ecological survey of Florida Bay estuaries financed by the State Board of Conservation, Marine Laboratory workers repeatedly captured many specimens of O. -

Characteristics of Estuarine Sediments of the United States
Characteristics of Estuarine Sediments of The United States GEOLOGICAL SURVEY PROFESSIONAL PAPER 742 Prepared in cooperation with the U.S. Federal Water Pollution Control Administration Characteristics of Estuarine Sediments of The United States By DAVID W. FOLGER GEOLOGICAL SURVEY PROFESSIONAL PAPER 742 Prepared in cooperation with the U.S. Federal Water Pollution Control Administration A compilation of data, essentially an atlas, on texture and composition of bottom sediments, including geologic and hydro logic factors that influence them, in 4.5 estuaries UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1972 UNITED STATES DEPARTMENT OF THE INTERIOR ROGERS C. B. MORTON, Secretary GEOLOGICAL SURVEY V. E. McKelvey, Director Library of Congress catalog-card No. 72-600206 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 - Price $1.25 (paper cover) Stock Number 2401-2112 CONTENTS Page Page Abstract_ ________________________ 1 Area summaries Continued Introduction _______________________ 1 St. Joseph Bay, Fla_______.- _. 44 Scope and purpose of the study_ 1 Pensacola Bay, Fla____--_-_-_--__-----------__- 46 Sources of information_________ 1 Mobile Bay, Ala___________________ _______ 48 Format______________________ 1 Mississippi Sound, Miss, and Ala_________________ 49 Setting.___________________ 3 Breton and Chandeleur Sounds, La_______________ 51 Sediment texture ___________ 3 Barataria Bay, La______________________________ 53 Sediment composition. ______ 3 Sabine Lake, Tex. and La______________________ -

Juvenile and Small Resident Fishes of Florida Bay, a Critical Habitat in The
NOAA Professional Paper NMFS 6 U.S. Department of Commerce March 2007 Juvenile and small resident fishes of Florida Bay, a critical habitat in the Everglades National Park, Florida Allyn B. Powell Gordon Thayer Michael Lacroix Robin Cheshire U.S. Department of Commerce NOAA Professional Carlos M. Gutierrez Secretary National Oceanic Papers NMFS and Atmospheric Administration Vice Admiral Scientific Editor Conrad C. Lautenbacher Jr., Dr. Adam Moles USN (ret.) Under Secretary for Technical Editor Oceans and Atmosphere Elizabeth Calvert National Marine Fisheries Service, NOAA National Marine 17109 Point Lena Loop Road Fisheries Service Juneau, AK 99801-8626 William T. Hogarth Assistant Administrator for Fisheries Managing Editor Shelley Arenas National Marine Fisheries Service Scientific Publications Office 7600 Sand Point Way NE Seattle, WA 98115 Editorial Committee Dr. Ann C. Matarese National Marine Fisheries Service Dr. James W. Orr National Marine Fisheries Service Dr. Bruce L. Wing National Marine Fisheries Service The NOAA Professional Paper NMFS (ISSN 1931-4590) series is published by the Scientific Publications Office, National Marine Fisheries Service, The NOAA Professional Paper NMFS series carries peer-reviewed, lengthy original NOAA, 7600 Sand Point Way NE, research reports, taxonomic keys, species synopses, flora and fauna studies, and data-in- Seattle, WA 98115. tensive reports on investigations in fishery science, engineering, and economics. Copies The Secretary of Commerce has of the NOAA Professional Paper NMFS series are available free in limited numbers to determined that the publication of government agencies, both federal and state. They are also available in exchange for this series is necessary in the transac- tion of the public business required by other scientific and technical publications in the marine sciences. -

Cooperative Gulf of Mexico Estuarine Inventory and Study, Florida / J
<-\^ C5 5.13 ; N^FS -3L'f NOAA TR NMFS CIRC-368 NOAA Technical Report NMFS CIRC-368 M,otc ^ °v U.S. DEPARTMENT OF COMMERCE National Oceanic and Atmospheric Administration \ :r National Marine Fisheries Service Cooperative Gulf of Mexico Estuarine Inventory and Study, Florida: Phase I, Area Description J. KNEELAND McNULTY, WILLIAM N. LINDALL, JR., AND JAMES E. SYKES SEATTLE, WA November 1972 NOAA TECHNICAL REPORTS National Marine Fisheries Service, Circulars The major responsibilities of the National Marine Fisheries Service (NMFS) are to monitor and assess the abundance and geographic distribution of fishery resources, to understand and predict fluctuations in the quan- tity and distribution of these resources, and to establish levels for optimum use of the resources. NMFS is also charged with the development and implementation of policies for managing national fishing grounds, develop- ment and enforcement of domestic fisheries regulations, surveillance of foreign fishing off United States coastal waters, and the development and enforcement of international fishery agreements and policies. NMFS also assists the fishi. g industry through marketing service and economic analysis programs, and mortgage insurance and vessel construction subsidies. It collects, analyses, and publishes statistics on various phases of the industry. The NOAA Technical Report NMFS CIRC series continues a series that has been in existence since 1941. The Circulars are technical publications of general interest intended to aid conservation and management. Publica- tions that review in considerable detail and at a high technical level certain broad areas of research appear in this series. Technical papers originating in economics studies and from management investigations appear in the Circular series. -

Proposed Methodology for the Assessment of Protective Numeric Nutrient Criteria for South Florida Estuaries and Coastal Waters
Proposed Methodology for the Assessment of Protective Numeric Nutrient Criteria for South Florida Estuaries and Coastal Waters Prepared by: Henry Briceño, Joseph Boyer and Peter Harlem Florida International University Southeast Environmental Research Center Miami, Florida Delivered to: U. S. Environmental Protection Agency Miami December 6th, 2010 This report is contribution # T-501 of the Southeast Environmental Research Center at Florida International University OE #148, Florida International University, Miami, Florida 33199 305‐348‐3095; 305‐348‐4096 fax; http://serc.fiu.edu Ms Stephanie Sanzone Designated Federal Officer(DFO) EPA Science Advisory Board Staff Office (1400R) U.S. Environmental Protection Agency 1200 Pennsylvania Avenue, NW Washington, DC 20460 Re: EPA SAB Nutrient Criteria Review Panel Dear Ms. Sanzone We would like to provide the attached document entitled “Proposed Methodology for the Assessment of Protective Nutrient Criteria for South Florida Estuaries and Coastal Waters” to the EPA Science Advisory Board, Nutrient Criteria Review Panel. We speak as members of South Florida scientific community, principal investigators for many years of a large marine water quality monitoring network, contributors of statistical and modeling approaches to the above document, and member (Boyer) of Florida’s Marine Numeric Nutrient Criteria Technical Advisory Committee (MTAC) Sincerely, Dr. Henry O. Briceño, Research Faculty Southeast Environmental Research Center Florida International University Dr. Joseph N. Boyer, Director and