Cyclic Distributional Model of the Late Moscovian Fusulinids in the Donets
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bryozoan Studies 2019
BRYOZOAN STUDIES 2019 Edited by Patrick Wyse Jackson & Kamil Zágoršek Czech Geological Survey 1 BRYOZOAN STUDIES 2019 2 Dedication This volume is dedicated with deep gratitude to Paul Taylor. Throughout his career Paul has worked at the Natural History Museum, London which he joined soon after completing post-doctoral studies in Swansea which in turn followed his completion of a PhD in Durham. Paul’s research interests are polymatic within the sphere of bryozoology – he has studied fossil bryozoans from all of the geological periods, and modern bryozoans from all oceanic basins. His interests include taxonomy, biodiversity, skeletal structure, ecology, evolution, history to name a few subject areas; in fact there are probably none in bryozoology that have not been the subject of his many publications. His office in the Natural History Museum quickly became a magnet for visiting bryozoological colleagues whom he always welcomed: he has always been highly encouraging of the research efforts of others, quick to collaborate, and generous with advice and information. A long-standing member of the International Bryozoology Association, Paul presided over the conference held in Boone in 2007. 3 BRYOZOAN STUDIES 2019 Contents Kamil Zágoršek and Patrick N. Wyse Jackson Foreword ...................................................................................................................................................... 6 Caroline J. Buttler and Paul D. Taylor Review of symbioses between bryozoans and primary and secondary occupants of gastropod -

Palaeontologia Electronica DISCRIMINATION OF
Palaeontologia Electronica http://palaeo-electronica.org DISCRIMINATION OF FENESTRATE BRYOZOAN GENERA IN MORPHOSPACE Steven J. Hageman and Frank K. McKinney ABSTRACT Concepts for generic diagnoses and discrimination of biserial fenestrate Bryozoa (Fenestellidae) have varied historically, but have largely been based on specialized colony forms (e.g., Archimedes), the shape and budding arrangement of chambers and other internal skeletal features such as hemisepta, and occasionally on the pres- ence or absence of discrete characters, such as placement of nodes on the frontal sur- face (e.g., Minilya). The question remains as to whether biserial fenestrate genera represent real biological clades, or whether they are convenient groupings of morpho- types based on untested characters. This study evaluates the distribution of 1075 operational taxonomic units (OTUs) from 15 fenestrate genera with measurements for nine morphometric characters – external features are not emphasized in most generic diagnoses. Here, each OTU represents a composite or idealized individual from a col- ony. Results show that OTUs plotted in principal component space do largely form coherent clusters based on a priori generic assignments. Thus the groupings based on characters other than the ones used to originally define them, add support to the notion of biological significance for recognized genera. The exceptions actually highlight and help resolve known issues. Therefore, we recognize Alternifenestella as a junior syn- onym of the genus Spinofenestella, and propose reassignment of Laxifenestella serrat- ula in Snyder (1991) to Fenestella serratula, and Fenestella sp. 1 in Ernst and Schroeder (2007) as Rectifenestella. We do not advocate that biserial fenestrate generic concepts should be based on the nine external characters used in this study, but rather that they can be used independently to evaluate the coherence of genera based on other discrete characters. -

The Carboniferous Evolution of Nova Scotia
Downloaded from http://sp.lyellcollection.org/ by guest on September 27, 2021 The Carboniferous evolution of Nova Scotia J. H. CALDER Nova Scotia Department of Natural Resources, PO Box 698, Halifax, Nova Scotia, Canada B3J 2T9 Abstract: Nova Scotia during the Carboniferous lay at the heart of palaeoequatorial Euramerica in a broadly intermontane palaeoequatorial setting, the Maritimes-West-European province; to the west rose the orographic barrier imposed by the Appalachian Mountains, and to the south and east the Mauritanide-Hercynide belt. The geological affinity of Nova Scotia to Europe, reflected in elements of the Carboniferous flora and fauna, was mirrored in the evolution of geological thought even before the epochal visits of Sir Charles Lyell. The Maritimes Basin of eastern Canada, born of the Acadian-Caledonian orogeny that witnessed the suture of Iapetus in the Devonian, and shaped thereafter by the inexorable closing of Gondwana and Laurasia, comprises a near complete stratal sequence as great as 12 km thick which spans the Middle Devonian to the Lower Permian. Across the southern Maritimes Basin, in northern Nova Scotia, deep depocentres developed en echelon adjacent to a transform platelet boundary between terranes of Avalon and Gondwanan affinity. The subsequent history of the basins can be summarized as distension and rifting attended by bimodal volcanism waning through the Dinantian, with marked transpression in the Namurian and subsequent persistence of transcurrent movement linking Variscan deformation with Mauritainide-Appalachian convergence and Alleghenian thrusting. This Mid- Carboniferous event is pivotal in the Carboniferous evolution of Nova Scotia. Rapid subsidence adjacent to transcurrent faults in the early Westphalian was succeeded by thermal sag in the later Westphalian and ultimately by basin inversion and unroofing after the early Permian as equatorial Pangaea finally assembled and subsequently rifted again in the Triassic. -

Influence of Different Factors on the Sediment Yield of the Oka Basin Rivers (Central Russia)
Sediment Dynamics and the Hydromorphology of Fluvial Systems (Proceedings of a symposium held in 28 Dundee, UK, July 2006). IAHS Publ. 306, 2006. Influence of different factors on the sediment yield of the Oka basin rivers (central Russia) VALENTIN GOLOSOV Laboratory for Soil Erosion and Fluvial Processes, Faculty of Geography, Moscow State University, GSP-2, 119992, Moscow, Russia [email protected] Abstract In order to investigate links between basin/land-use characteristics and sediment fluxes of rivers within the Oka River basin, a database has been compiled from observations obtained at 25 gauging stations located in different parts of the basin. Relatively high correlations have been found between sediment yield from basin hillslopes and river sediment yield for: (i) rivers of the forest and northern part of forest-steppe zones (r2 = 0.50); and (ii) rivers of forest-steppe zones (r2 = 0.52). A linear positive relationship (r2 = 0.71) has been found between sediment delivery ratio and weighted average gradient of river channel separately for the large river basins and small river basins of the forest zone. A negative relationship between sediment delivery ratio and forested area within basins is found separately for the small (S < 2500 km2) rivers and large rivers of the Oka River basin. The influence of other factors as well as the problem of small river aggradation is also discussed. Key words erosion; gauging station; river aggradation; Russia; sediment yield INTRODUCTION Under natural conditions, the contribution of the basin-derived sediment yield component decreases and the channel-derived component increases slowly from the upper parts of fluvial systems to the large river basins (Dedkov & Mozzerin, 2000). -

Mississippian and Pennsylvanian Fossils of the Albuquerque Country Stuart A
New Mexico Geological Society Downloaded from: http://nmgs.nmt.edu/publications/guidebooks/12 Mississippian and Pennsylvanian fossils of the Albuquerque country Stuart A. Northrop, 1961, pp. 105-112 in: Albuquerque Country, Northrop, S. A.; [ed.], New Mexico Geological Society 12th Annual Fall Field Conference Guidebook, 199 p. This is one of many related papers that were included in the 1961 NMGS Fall Field Conference Guidebook. Annual NMGS Fall Field Conference Guidebooks Every fall since 1950, the New Mexico Geological Society (NMGS) has held an annual Fall Field Conference that explores some region of New Mexico (or surrounding states). Always well attended, these conferences provide a guidebook to participants. Besides detailed road logs, the guidebooks contain many well written, edited, and peer-reviewed geoscience papers. These books have set the national standard for geologic guidebooks and are an essential geologic reference for anyone working in or around New Mexico. Free Downloads NMGS has decided to make peer-reviewed papers from our Fall Field Conference guidebooks available for free download. Non-members will have access to guidebook papers two years after publication. Members have access to all papers. This is in keeping with our mission of promoting interest, research, and cooperation regarding geology in New Mexico. However, guidebook sales represent a significant proportion of our operating budget. Therefore, only research papers are available for download. Road logs, mini-papers, maps, stratigraphic charts, and other selected content are available only in the printed guidebooks. Copyright Information Publications of the New Mexico Geological Society, printed and electronic, are protected by the copyright laws of the United States. -
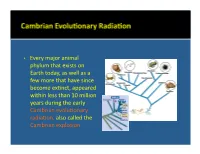
• Every Major Animal Phylum That Exists on Earth Today, As Well As A
• Every major animal phylum that exists on Earth today, as well as a few more that have since become ex:nct, appeared within less than 10 million years during the early Cambrian evolu:onary radiaon, also called the Cambrian explosion. • Phylum Pro:sta includes a diverse group of eukaryo:c microorganisms which are mostly unicellular. They are not necessarily primi:ve, although some are. Only some have a skeleton that can be preserved as a fossil. • Foraminifera and radiolaria are the most important examples. Coccolithophores, diatoms, and dinoflagellates are also examples. • Foraminifera are marine organisms that may be either planktonic, living in the water column where they float at various levels, or benthic, living on or within the seafloor sediment. • They form relavely large (0.1 mm to 5 cm) porous internal skeletons called tests through which project strands of living cytoplasm. The single- to mul:-chambered tests may be composed of organic maer, agglu:nated sand grains or sponge spicules, or calcium carbonate. • Benthic foraminifera • Planktonic foraminifera (Cambrian to Recent). (Jurassic to Recent). Oligocene Eocene • Radiolaria are planktonic marine organisms that tend to live in relavely cold water. • Their tests, oNen delicate and elaborate, are made of silica and presently accumulate on the floors of the parts of oceans that are deeper than those where foraminiferal tests are accumulang. • Radiolaria (Cambrian to • Radiolaria (Cambrian to Recent). Recent). Paleocene Radiolaria • Coccolithophores (Triassic to Recent) are extremely small marine planktonic photosynthe:c unicellular algae that are enclosed by plates of low-Mg calcite called coccoliths. Cretaceous White Cliffs of Dover, England • Diatoms (Jurassic to Recent) are small freshwater and marine planktonic photosynthe:c unicellular algae that are enclosed by a cell wall made of silica called a frustule. -

Material Composition and Geochemical Characteristics of Technogenic River Silts E
ISSN 0016-7029, Geochemistry International, 2019, Vol. 57, No. 13, pp. 1361–1454. © Pleiades Publishing, Ltd., 2019. Material Composition and Geochemical Characteristics of Technogenic River Silts E. P. Yanin* Vernadsky Institute of Geochemistry and Analytical Chemistry, Russian Academy of Sciences, Moscow, 119991 Russia *e-mail: [email protected] Received May 20, 2019; revised June 7, 2019; accepted June 7, 2019 Abstract—The paper discusses the results of many years of studying the material composition and geochem- ical characteristics, conditions, and processes in the formation of technogenic river silts: a new type of mod- ern river sediments formed in riverbeds within the boundaries and zones of influence of industrial–urbanized areas. The article examines the main sources and most important characteristics of technogenic sedimentary material flowing into rivers, as well as the geochemical conditions of technogenic alluvial sedimentation, the morphology and structure of technogenic silts, the extent of their spatial distribution in riverbeds, their grain size characteristics, and mineral and chemical composition. Special attention is paid to analyzing the group composition of organic matter in river sediments and the features of its transformation in pollution zones. The study analyzes the technogenic geochemical associations that form in silts in zones of influence of various impact sources, the features of the concentration and distribution of chemical elements, heavy metal specia- tion, the composition of exchangeable cations in technogenic silts and natural (background) alluvium, and the composition of silt water. Possible secondary transformations of technogenic silts and their significance as a long-term source of pollution of the water mass and hydrobionts are substantiated. -

Fenestrate Bryozoans from the Glen Dean Limestone (Middle Chester) of Southern Indiana
FENESTRATE BRYOZOANS FROM THE GLEN DEAN LIMESTONE (MIDDLE CHESTER) OF SOUTHERN INDIANA by JOHN UTGAARD T. G. PERRY Indiana Department of Conservation GEOLOGICAL SURVEY Bulletin No. 19 1960 STATE OF INDIANA HAROLD W. HANDLEY, GOVERNOR DEPARTMENT OF CONSERVATION E. KENNETH MARLIN, DIRECTOR GEOLOGICAL SURVEY JOHN B. PATTON, STATE GEOLOGIST BLOOMINGTON ___________________________________________________________________________________________ BULLETIN NO. 19 ___________________________________________________________________________________________ FENESTRATE BRYOZOANS FROM THE GLEN DEAN LIMESTONE (MIDDLE CHESTER) OF SOUTHERN INDIANA BY JOHN UTGAARD T. G. PERRY PRINTED BY AUTHORITY OF THE STATE OF INDIANA BLOOMINGTON, INDIANA December 1960 ___________________________________________________________________________________________ For sale by Geological Survey, Indiana Department of Conservation, Bloomington, Ind. Price $1.25 2 SCIENTIFIC AND TECHNICAL STAFF OF THE GEOLOGICAL SURVEY JOHN B. PATTON, State Geologist MAURICE E. BIGGS, Assistant State Geologist MARY BETH FOX, Mineral Statistician Coal Section Charles E. Wier, Geologist and Head G. K. Guennel, Paleobotanist S. A. Friedman, Geologist Harold C. Hutchison, Geologist Richard C. Neavel, Coal Petrographer William C. Richardson, Geological Assistant Drafting and Photography Section William H. Moran, Chief Draftsman Robert E. Judah, Geological Artist-Draftsman Micky P. Love, Geological Draftsman John E. Peace, Senior Geological Draftsman George R. Ringer, Photographer Educational Services R. Dee Rarick, Geologist and Head Geochemis try Section R. K. Leininger, Spectrographer and Head Maynard E. Coller, Chemist Louis V. Miller, Coal Chemist E. M. Craig, Geochemical Assistant Geology Section Robert H. Shaver, Paleontologist and Head Henry H. Gray, Head Stratigrapher William J. Wayne, Head Glacial Geologist Allan F. Schneider, Glacial Geologist Geophysics Section Maurice E. Biggs, Geophysicist and Head Robert F. Blakely, Geophysicist Charles S. Miller, Instrument Maker Albert J. Rudman, Geophysicist Joseph F. -

Chapter 5. Paleozoic Invertebrate Paleontology of Grand Canyon National Park
Chapter 5. Paleozoic Invertebrate Paleontology of Grand Canyon National Park By Linda Sue Lassiter1, Justin S. Tweet2, Frederick A. Sundberg3, John R. Foster4, and P. J. Bergman5 1Northern Arizona University Department of Biological Sciences Flagstaff, Arizona 2National Park Service 9149 79th Street S. Cottage Grove, Minnesota 55016 3Museum of Northern Arizona Research Associate Flagstaff, Arizona 4Utah Field House of Natural History State Park Museum Vernal, Utah 5Northern Arizona University Flagstaff, Arizona Introduction As impressive as the Grand Canyon is to any observer from the rim, the river, or even from space, these cliffs and slopes are much more than an array of colors above the serpentine majesty of the Colorado River. The erosive forces of the Colorado River and feeder streams took millions of years to carve more than 290 million years of Paleozoic Era rocks. These exposures of Paleozoic Era sediments constitute 85% of the almost 5,000 km2 (1,903 mi2) of the Grand Canyon National Park (GRCA) and reveal important chronologic information on marine paleoecologies of the past. This expanse of both spatial and temporal coverage is unrivaled anywhere else on our planet. While many visitors stand on the rim and peer down into the abyss of the carved canyon depths, few realize that they are also staring at the history of life from almost 520 million years ago (Ma) where the Paleozoic rocks cover the great unconformity (Karlstrom et al. 2018) to 270 Ma at the top (Sorauf and Billingsley 1991). The Paleozoic rocks visible from the South Rim Visitors Center, are mostly from marine and some fluvial sediment deposits (Figure 5-1). -

«Faymonville »
65 13/104 Июль 2020 With the support of the CCI of Russia 12+ ОЛЕГ ПРОЗОРОВ: !СИНЕРГИЯ ЧЛЕНОВ БЕЛЬГИЙСКО1 ЛЮКСЕМБУРГСКОЙ ТОРГОВОЙ ПАЛАТЫ В РОССИИ 9 ДЕЙСТВОВАТЬ И ПРЕУСПЕВАТЬ СООБЩА< НА НОВЫЕ ЛЬГОТЫ ДЛЯ КАЛИНИНГРАДСКОГО БИЗНЕСА ВЫДЕЛИЛИ БОЛЕЕ 200 МЛН РУБЛЕЙ ДМИТРИЙ КОБЫЛКИН: 7РАЗВИТИЕ ЭКОЛОГИИ 9 ЗАЛОГ ЗДОРОВОЙ ЖИЗНИ ГРАЖДАН СТРАНЫ; ЕЛЕНА ФАДЕЕВА, генеральный директор «ФЭЙМОНВИЛЬ РУСЬ»: «FAYMONVILLE ЯВЛЯЕТСЯ ВЕДУЩИМ МИРОВЫМ ПРОИЗВОДИТЕЛЕМ СПЕЦИАЛЬНЫХ ТРАНСПОРТНЫХ СРЕДСТВ ДЛЯ ПЕРЕВОЗКИ ТЯЖЁЛЫХ ГРУЗОВ» 66 RUSSIAN BUSINESS GUIDE =ИЮЛЬ 2020> CONTENTS | СОДЕРЖАНИЕ 1 Russian Business Guide ОФИЦИАЛЬНО. OFFICIALLY www.rbgmedia.ru ОЛЕГ ПРОЗОРОВ: «СИНЕРГИЯ ЧЛЕНОВ БЕЛЬГИЙСКО-ЛЮКСЕМБУРГСКОЙ Деловое издание, рассказывающее о развитии, 2 ТОРГОВОЙ ПАЛАТЫ В РОССИИ – ДЕЙСТВОВАТЬ И ПРЕУСПЕВАТЬ СООБЩА» отраслях, перспективах, персоналиях бизнеса в России и за рубежом. OLEG PROZOROV: "SYNERGY OF THE MEMBERS OF THE BELGIAN-LUXEMBOURG 12+ CHAMBER OF COMMERCE IN RUSSIA – TO ACT AND SUCCEED TOGETHER" Учредитель и издатель: ООО «БИЗНЕС-ДИАЛОГ МЕДИА» ЛИЦО С ОБЛОЖКИ. COVER STORY при поддержке ТПП РФ ГРУППА КОМПАНИЙ FAYMONVILLE: КАЧЕСТВО, ПРОФЕССИОНАЛИЗМ, Редакционная группа: 6 НАДЁЖНОСТЬ Максим Фатеев, Вадим Винокуров, Наталья Чернышова FAYMONVILLE GROUP: QUALITY, PROFESSIONALISM, RELIABILITY Главный редактор: Мария Сергеевна Суворовская Редактор номера: ОТ ПЕРВОГО ЛИЦА. IN THE FIRST PERSON София Антоновна Коршунова ЗДЕСЬ УМЕЮТ СТРОИТЬ КОРАБЛИ Заместитель директора по коммерческим вопросам: 14 PEOPLE HERE KNOW HOW TO BUILD SHIPS Ирина Владимировна Длугач Дизайн/вёрстка: Александр Лобов Перевод: Мария Ключко АЛЕКСАНДР АНИКЕЕВ, АО «ЭКОПЭТ»: «НАШ ЗАВОД НА СЕГОДНЯ – Отпечатано в типографии ООО «ВИВА-СТАР», 20 САМОЕ ИННОВАЦИОННОЕ ПРЕДПРИЯТИЕ В МИРЕ!» г. Москва, ул. Электрозаводская, д. 20, стр. 3. ALEXANDER ANIKEEV, JSC EKOPET: “OUR PLANT IS THE MOST INNOVATIVE Материалы, отмеченные значком R или «РЕКЛАМА», публикуются на правах рекламы. -

Departure City City Of Delivery Region Delivery Delivery Time
Cost of Estimated Departure city city of delivery Region delivery delivery time Moscow Ababurovo Moscow 655 1 Moscow Abaza The Republic of Khakassia 1401 6 Moscow Abakan The Republic of Khakassia 722 2 Moscow Abbakumova Moscow region 655 1 Moscow Abdrakhmanovo Republic of Tatarstan 682 on request Moscow Abdreevo Ulyanovsk region 1360 5 Moscow Abdulov Ulyanovsk region 1360 5 Moscow Abinsk Krasnodar region 682 3 Moscow Abramovka Ulyanovsk region 1360 5 Moscow Abramovskikh Sverdlovsk region 1360 1 Moscow Abramtsevo Moscow region 655 1 Moscow Abramtzevo (Dmitrovsky reg) Moscow region 1360 3 Moscow Abrau Durso Krasnodar region 682 1 Moscow Avvakumova Tver region 655 5 Moscow Avdotyino Moscow region 655 1 Moscow Avdotyino (Stupinsky reg) Moscow region 1360 1 Averkieva Moscow Moscow region 1360 2 (Pavlovsky Posadskiy reg) Aviation workers Moscow Moscow region 1360 1 (Odintsovskiy-one) Moscow aviators Moscow region 655 1 Moscow Aviation Moscow region 655 1 Moscow Aviation Moscow region 655 1 Moscow Motorist Arhangelsk region 655 1 Moscow avtopoligone Moscow region 1360 3 Moscow Autoroute Moscow region 655 1 Moscow agarin Moscow region 655 1 Moscow Agarin (Stupinsky reg) Moscow region 1360 1 Moscow Agafonov Moscow region 655 1 Moscow AGAFONOVA (Odintsovskiy-one) Moscow region 1360 1 Moscow Agashkino Moscow region 655 5 Moscow Ageevka Oryol Region 655 1 Moscow Agidel Republic of Bashkortostan 1360 3 Moscow Agha Krasnodar region 682 3 Moscow Agrarnik Tver region 1306 6 Moscow agricultural Republic of Crimea 682 4 Moscow agrogorodok Moscow region -

Dimensional DELFT-3D Model, the Case Study of the Oka River
E3S Web of Conferences 163, 01012 (2020) https://doi.org/10.1051/e3sconf/202016301012 IV Vinogradov Conference Maximum water level calculation based on 2- dimensional DELFT-3D model, the case study of the Oka river Pavel Terskii1,2*, Elizaveta Rakcheeva2, Maksim Kharlamov1, Andrey Pavlovskiy2, and Igor Zemlyanov2 1 Lomonosov Moscow State University, 1 Leninskie Gory, 119991, Moscow, Russia 2 Zubov State Oceanographic Institute, 6 Kropotkinskiy lane, 119034, Moscow, Russia Abstract. This research represents methodical approach and main results of water level hydrodynamic modeling for real summer low-water season and extreme spring flood of 1% probability. The object of modeling is 105 km length section of the Oka River in Moscow region between Kashira and Kolomna urban districts. Modeling object includes also the Moscow River section downstream the Severka River mouth. Two-dimensional hydrodynamic model constructed using DELFT-3D. To build the model we used the detailed elevation model and hydrological conditions based on in-situ measurements in 2019, special estimated discharge and water level time-series of 1% exceedance probability based on reference year observations and distributed Chezy coefficient calibration. The model implementation resulted in the actual detailed distribution of the water discharge, velocity and level along the Oka River section (including the downstream Moscow River) for low-water and extremely high water periods. These results are used for the flood zone delineation in the Kashira and Kolomna urban districts. 1 Introduction Hydrological processes may cause economic losses and social risk in some cases of their dangerous natural conditions. These days Russian Federation has a relatively new regulation – the Government Decision #360 on the Flood zones delineation and regulation.