Founding, Foraging, and Fighting: Colony Size and the Spatial Distribution of Harvester Ant Nests Author(S): Deborah M
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Double Mound of the Harvester Ant Pogonomyrmex Occidentalis (Hymenoptera: Formicidae, Myrmicinae)
Great Basin Naturalist Volume 53 Number 4 Article 13 12-28-1993 Double mound of the harvester ant Pogonomyrmex occidentalis (Hymenoptera: Formicidae, Myrmicinae) William H. Clark Orma J. Smith Museum of Natural History, Albertson College of Idaho, Caldwell, Idaho Follow this and additional works at: https://scholarsarchive.byu.edu/gbn Recommended Citation Clark, William H. (1993) "Double mound of the harvester ant Pogonomyrmex occidentalis (Hymenoptera: Formicidae, Myrmicinae)," Great Basin Naturalist: Vol. 53 : No. 4 , Article 13. Available at: https://scholarsarchive.byu.edu/gbn/vol53/iss4/13 This Note is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Great Basin Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Creat Basin Naturalist 53(4), pp. 407-408 DOUBLE MOUND OF THE HARVESTER ANT POGONOMYRMEX OCCLDENTALIS (HYMENOPTERA: FORMICIDAE, MYRMICINAE) William H. ClarkI Key words: ho.rvester ant double mound, Pogonomyrmex occidentalis, ant nest, UUilt. Several species ofPogOlWmynnex, especial County, Utah, approximately 58 Ian NE Moab ly members of the occidentalis complex, have along Highway 128 and about 4 Ian from the nests surmounted by a large, conical mound Colorado River, at an elevation of 1445 m. of soil and gravel in a clearing created by the Voucher specimens from both mounds (WHC ants (Cole 1932, 1968). I report a rare double #8184 and 8185) are deposited in the Orma J. mound of the western halvester ant, Pogono Smith Museum of Natural History, Albertson mymwx oecidentalis (Cresson), having two College ofIdaho, Caldwell (ClDA). -

The Ants of Oklahoma Master of Science
THE ANTS OF OKLAHOMA By Jerry H. Young(I\" Bachelor of Science Oklahoma Agricultural and Mechanical College Stillwater, Oklahoma 1955 Submitted to the faculty of the Graduate School of the Oklahoma Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE January 1 1956 tl<lAWMA AGCMCl«.f�Al L �Ci'!AlttCAl e&U.Ull LIBRARY JUL16195 6 THE ANTS OF OKLAHOMA Thesis Approved: Thesis Adviser }>JcMem��f � 't'" he Thesis ) Committee Member of the Thesis Committee 7'4'.��Member of the Thesis Committee Head of the Department ifean of the Graduate School 361565 ii PREFACE The study of the distribution of ants in the United States has been a long and continuous process with many contributors, but the State of Oklahoma has not received the attentions of these observers to any great extent. The only known list of ants of Oklahoma is one prepared by Mo Ro Smith (1935)0 Early in 1954 a survey of the state of Oklahoma was made to determine the species present and their distributiono The results of this survey, which blanketed the entire State, are given in this paper. The author wishes to express his appreciation to Dro Do E. Howell, chairman of the writer's thesis committee, for his valuable assistance and careful guidance in the preparation of this papero Also, much guidance on preparation of this manuscrip_t was received from Drs. Do Eo Bryan, William H. Irwin and F. A. Fenton. Many of the determin ations were made by M. R. Smith.. Vital infonnation was obtained from the museums at Oklahoma Agricultural and Mechanical College and the University of Oklahoma. -

Wildland Fire in Ecosystems: Effects of Fire on Fauna
United States Department of Agriculture Wildland Fire in Forest Service Rocky Mountain Ecosystems Research Station General Technical Report RMRS-GTR-42- volume 1 Effects of Fire on Fauna January 2000 Abstract _____________________________________ Smith, Jane Kapler, ed. 2000. Wildland fire in ecosystems: effects of fire on fauna. Gen. Tech. Rep. RMRS-GTR-42-vol. 1. Ogden, UT: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. 83 p. Fires affect animals mainly through effects on their habitat. Fires often cause short-term increases in wildlife foods that contribute to increases in populations of some animals. These increases are moderated by the animals’ ability to thrive in the altered, often simplified, structure of the postfire environment. The extent of fire effects on animal communities generally depends on the extent of change in habitat structure and species composition caused by fire. Stand-replacement fires usually cause greater changes in the faunal communities of forests than in those of grasslands. Within forests, stand- replacement fires usually alter the animal community more dramatically than understory fires. Animal species are adapted to survive the pattern of fire frequency, season, size, severity, and uniformity that characterized their habitat in presettlement times. When fire frequency increases or decreases substantially or fire severity changes from presettlement patterns, habitat for many animal species declines. Keywords: fire effects, fire management, fire regime, habitat, succession, wildlife The volumes in “The Rainbow Series” will be published during the year 2000. To order, check the box or boxes below, fill in the address form, and send to the mailing address listed below. -

Influence of Small-Scale Disturbances by Kangaroo Rats on Chihuahuan Desert Ants
Oecologia (2000) 125:142–149 © Springer-Verlag 2000 R.L. Schooley · B.T. Bestelmeyer · J.F. Kelly Influence of small-scale disturbances by kangaroo rats on Chihuahuan Desert ants Received: 11 October 1999 / Accepted: 11 March 2000 Abstract Banner-tailed kangaroo rats (Dipodomys Introduction spectabilis) are prominent ecosystem engineers that build large mounds that influence the spatial structuring Natural disturbances create environmental patchiness of fungi, plants, and some ground-dwelling animals. that can strongly influence organisms and ecological Ants are diverse and functionally important components processes (Watt 1947; Wiens 1976; Pickett and White of arid ecosystems; some species are also ecosystem en- 1985; Turner 1987). In particular, patch disturbances can gineers. We investigated the effects of patch disturbances affect patterns of species richness and community struc- created by D. spectabilis mounds on ant assemblages in ture by increasing habitat heterogeneity and permitting a Chihuahuan Desert grassland in southern New Mexico the coexistence of species with differing competitive and by using pitfall traps in a paired design (mound vs. ma- colonization abilities (Horn and MacArthur 1972; trix). Although the disturbances did not alter species Denslow 1985; Huston 1994). In terrestrial systems, the richness or harbor unique ant communities relative to the disturbances created by the activities of burrowing mam- matrix, they did alter species composition; the abun- mals are important generators of spatial heterogeneity dances of 6 of 26 species were affected. The distur- (Huntley and Inouye 1988; Whicker and Detling 1988; bances might also act to disrupt spatial patterning of ants Hansell 1993). These effects are especially evident in ar- caused by other environmental gradients. -

Phrynosoma Cornutum and P
STRATEGIES OF PREDATORS AND THEIR PREY: OPTIMAL FORAGING AND HOME RANGE BEHAVIOR OF HORNED LIZARDS (PHRYNOSOMA SPP.) AND RESPONSE BY HARVESTER ANTS (POGONOMYRMEX DESERTORUM). Item Type text; Dissertation-Reproduction (electronic) Authors MUNGER, JAMES CAMERON. Publisher The University of Arizona. Rights Copyright © is held by the author. Digital access to this material is made possible by the University Libraries, University of Arizona. Further transmission, reproduction or presentation (such as public display or performance) of protected items is prohibited except with permission of the author. Download date 10/10/2021 20:56:51 Link to Item http://hdl.handle.net/10150/185315 INFORMATION TO USERS This reproduction was made from a copy of a document sent to us for microfilming. While the most advanced technology has been used to photograph and reproduce this document, the quality of the reproduction is heavily dependent upon the quality of the material submitted. The following explanation of techniques is provided to help clarify markings or notations which may appear on this reproduction. 1. The sign or "target" for pages apparently lacking from the document photographed is "Missing Page(s)". If it was possible to obtain the missing page(s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting through an image and duplicating adjacent pages to assure complete continuity. 2. When an image on the film is obliterated with a round black mark, it is an indication of either blurred copy because of movement during exposure, duplicate copy, or copyrighted materials that should not have been filmed. For blurred pages, a good image of the page can be found in the adjacent frame. -

Honey Bee (Apis Mellifera) Intracolonial Genetic Diversity Influences Worker Nutritional Status Bruce J
Honey bee (Apis mellifera) intracolonial genetic diversity influences worker nutritional status Bruce J. Eckholm, Ming H. Huang, Kirk E. Anderson, Brendon M. Mott, Gloria Degrandi-Hoffman To cite this version: Bruce J. Eckholm, Ming H. Huang, Kirk E. Anderson, Brendon M. Mott, Gloria Degrandi-Hoffman. Honey bee (Apis mellifera) intracolonial genetic diversity influences worker nutritional status. Api- dologie, Springer Verlag, 2015, 46 (2), pp.150-163. 10.1007/s13592-014-0311-4. hal-01284439 HAL Id: hal-01284439 https://hal.archives-ouvertes.fr/hal-01284439 Submitted on 7 Mar 2016 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Apidologie (2015) 46:150–163 Original article * INRA, DIB and Springer-Verlag France, 2014 DOI: 10.1007/s13592-014-0311-4 Honey bee (Apis mellifera ) intracolonial genetic diversity influences worker nutritional status 1 2 3 3 Bruce J. ECKHOLM , Ming H. HUANG , Kirk E. ANDERSON , Brendon M. MOTT , 3 Gloria DEGRANDI-HOFFMAN 11025 Zylstra Road, Coupeville, WA 98239, USA 2Eurofins Agroscience Services, Inc., 15250 NC Hwy 86 South, Prospect Hill, NC 27314, USA 3Carl Hayden Bee Research Center, USDA-ARS, 2000 East Allen Road, Tucson, AZ 85719, USA Received 9 January 2014 – Revised 10 July 2014 – Accepted 25 July 2014 Abstract – Honey bee queens mate with multiple males resulting in high intracolonial genetic diversity among nestmates; a reproductive strategy known as extreme polyandry. -

Pogonomyrmex Barbatus
Anita. Behav., 1986, 34, 1402-1419 The dynamics of the daily round of the harvester ant colony (Pogonomyrmex barbatus) DEBORAH M. GORDON Museum of Comparative Zoology and Society of Fellows, Harvard University, Cambridge, Massachusetts 02138, U.S.A. Abstract. Colonies of the red harvester ant, Pogonomyrmex barbatus, do various tasks outside the nest. There is a daily temporal pattern in the numbers of ants engaged in each of five activities: foraging, nest maintenance, patrolling, midden work and convening. Perturbations were carried out in the field to investigate how the daily round changes in response to environmental events and colony needs. Interfering with nest maintenance, foraging or both caused changes in the temporal patterns in all five of the observed activities. Removing nest maintenance workers, foragers or both caused the numbers involved in all five activities to decrease, and there were temporal patterns in the effects of removals. The results of both interference and removal experiments show that the extent to which a worker group does one activity affects the behaviour of other groups. When nest maintenance or foraging is impeded experimentally, these two activities are of reciprocal priority. When both are impeded, foraging is of higher priority than nest maintenance. Harvester ants forage, patrol, maintain the nest speaking, that of an equilibrium state, but the area and foraging trails, collect and arrange dynamics of colony organization are not yet under- pebbles on the nest, and gather in small groups, stood. What events will alter the daily round, by inspecting and grooming each other. The beha- how much, and for how long? Is there more than viour of the colony outside the nest at any moment one equilibrium state? can be described by citing the number of ants This study begins an exploration of the dynamics engaged in each of the five activities. -

Evolution of Colony Characteristics in the Harvester Ant Genus
Evolution of Colony Characteristics in The Harvester Ant Genus Pogonomyrmex Dissertation zur Erlangung des naturwissenschaftlichen Doktorgrades der Bayerischen Julius-Maximilians-Universität Würzburg vorgelegt von Christoph Strehl Nürnberg Würzburg 2005 - 2 - - 3 - Eingereicht am: ......................................................................................................... Mitglieder der Prüfungskommission: Vorsitzender: ............................................................................................................. Gutachter : ................................................................................................................. Gutachter : ................................................................................................................. Tag des Promotionskolloquiums: .............................................................................. Doktorurkunde ausgehändigt am: ............................................................................. - 4 - - 5 - 1. Index 1. Index................................................................................................................. 5 2. General Introduction and Thesis Outline....................................................... 7 1.1 The characteristics of an ant colony...................................................... 8 1.2 Relatedness as a major component driving the evolution of colony characteristics.................................................................................................10 1.3 The evolution -

Arizona Wildlife Notebook
ARIZONA WILDLIFE CONSERVATION ARIZONA WILDLIFE NOTEBOOK GARRY ROGERS Praise for Arizona Wildlife Notebook “Arizona Wildlife Notebook” by Garry Rogers is a comprehensive checklist of wildlife species existing in the State of Arizona. This notebook provides a brief description for each of eleven (11) groups of wildlife, conservation status of all extant species within that group in Arizona, alphabetical listing of species by common name, scientific names, and room for notes. “The Notebook is a statewide checklist, intended for use by wildlife watchers all over the state. As various individuals keep track of their personal observations of wildlife in their specific locality, the result will be a more selective checklist specific to that locale. Such information would be vitally useful to the State Wildlife Conservation Department, as well as to other local agencies and private wildlife watching groups. “This is a very well-documented snapshot of the status of wildlife species – from bugs to bats – in the State of Arizona. Much of it should be relevant to neighboring states, as well, with a bit of fine-tuning to accommodate additions and deletions to the list. “As a retired Wildlife Biologist, I have to say Rogers’ book is perhaps the simplest to understand, yet most comprehensive in terms of factual information, that I have ever had occasion to peruse. This book should become the default checklist for Arizona’s various state, federal and local conservation agencies, and the basis for developing accurate local inventories by private enthusiasts as well as public agencies. "Arizona Wildlife Notebook" provides a superb starting point for neighboring states who may wish to emulate Garry Rogers’ excellent handiwork. -

TAXONOMIC IDENTITY of "HALLUCINOGENIC" HARVESTER ANT Lpogonomyrmex Calijornicus) CONFIRMED
Journal of Ethnobiology 21(2): 133-144 Winter 2001 TAXONOMIC IDENTITY OF "HALLUCINOGENIC" HARVESTER ANT lPogonomyrmex calijornicus) CONFIRMED KEVIN I' GROARK Department of Anthropology, Un;ocrsity of California, Los Angeles Los Angeles, CA 90024 ABSTRACf.-The use of California harvester ants (fbgollomyrmex californiClls) for visionary and therapeutic ends was an important but poorly-documented tradi tion in native south-central California. In this brief report, a confirmation of the taxonomic identity of the red ant species used in California is presented, and the descriptive record of its use is supplemented with additional ethnographic ac counts. This taxonomic identification of this species is of particular importance, as visionary red ant ingestion provides the only well-documented case of the widespread use of an insect as an hallucinogenic agent. RESUMEN.-La utilizaci6n de hormigas granivoras rojas (HJgo/wmyrmex califor nicus) con fines alucin6genos y terapeuticos, fue una tradici6n de mucha impor tancia pero mal documentada en el sur y centro-sur de California. Este breve articulo confirma la identidad taxon6mica de dicha especie y la descripci6n de su uso se hace a !raves de datos etnogrMicos adicionales. Esta identificaci6n taxo n6mica es de especial interes, puesto que es el dnico ejemplo etnografico debi damente documentado de un agente alucin6geno derivado de un insecto. REsUME.-L'u!ilisation des fourmis moissonneuses rouges (fbgonomyrmex califor nicus) a des desseins religieux et therapeutiques etait une tradition peu docu mentee mais importante dans la vie de plusieurs groupes autochtones du centre sud de la Californie. Dans ce bref expose on trouve la confirmation de I'identification taxonomique de la fourmi et a la description de la methode de son utilisation s'ajoute des donnees ethnographiques suplementaires. -

Red Harvester Ant (Big Red Ants; Harvester Ants, Red Ants; Barbatus Harvester Ant)
Pest Profile Photo credit: April Nobile, California Academy of Sciences (Specimen CASENT0006306; from https://www.antweb.org) Common Name: Red harvester ant (big red ants; harvester ants, red ants; Barbatus harvester ant) Scientific Name: Pogonomyrmex barbatus Order and Family: Order Hymenoptera; Family Formicidae Size and Appearance: Length (mm) Appearance Egg Larva/Nymph Adult Workers Pogonomyrmex harvester ants are characterized by broad, range from 5 boxy heads, 12-segmented antennae, a two-part waist, a pair mm – 8 mm of dorsal spines and a stinger. Many species have a set of long hairs (called a psammophore) on the posterior lower portion of the head, behind the mouthparts. Workers of the red harvester ant are polymorphic without distinct majors and minors. They vary in color from a light to very deep red and they have texture in the form of lines on the head (rugae). Colonies have one queen (monogynous). Queens are similar to workers but larger with a larger thorax. Female and male reproductive ants have wings. Pupa (if applicable) Type of feeder (Chewing, sucking, etc.): Chewing Host(s): Harvester ants in the genus Pogonomyrmex specialize in seeds but may forage on other foods (generalist and opportunistic). Description of Damage (larvae and adults): Because theses ants primarily eat seeds (up to 90% of the diet), they are pests of agricultural systems, damaging crops but especially grasslands (pasture). They harvest seeds, defoliate plants, and remove young and old plants. Agricultural important plants they affect include corn, oats, alfalfa, cotton, guayule, grapes, date, citrus trees, apple, and pear, as well as more generally pasture (grasses) and shrubs. -
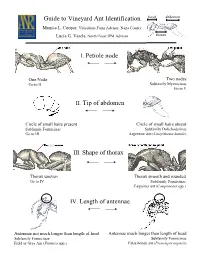
I. Petiole Node II. Tip of Abdomen IV. Length of Antennae Guide to Vineyard Ant Identification III. Shape of Thorax
Guide to Vineyard Ant Identification head abdomen Monica L. Cooper, Viticulture Farm Advisor, Napa County Lucia G. Varela, North Coast IPM Advisor thorax I. Petiole node One Node Two nodes Go to II Subfamily Myrmicinae Go to V II. Tip of abdomen Circle of small hairs present Circle of small hairs absent Subfamily Formicinae Subfamily Dolichoderinae Go to III Argentine Ant (Linepithema humile) III. Shape of thorax Thorax uneven Thorax smooth and rounded Go to IV Subfamily Formicinae Carpenter ant (Camponotus spp.) IV. Length of antennae Antennae not much longer than length of head Antennae much longer than length of head Subfamily Formicinae Subfamily Formicinae Field or Gray Ant (Formica spp.) False honey ant (Prenolepis imparis) head abdomen Petiole with two nodes Subfamily Myrmicinae (V-VIII) thorax V. Dorsal side of Thorax & Antennae One pair of spines on thorax No spines on thorax 12 segmented antennae 10 segmented antennae Go to VI Solenopsis molesta and Solenopsis xyloni VI. Underside of head No brush of bristles Brush of long bristles Go to VII Harvester ants (Pogonomyrmex californicus and P. brevispinosis) VII. Head and Thorax With hairs Without hairs Go to VIII Cardiocondyla mauritanica VIII. Head and Thorax With many parallel furrows Without parallel furrows Profile of thorax rounded Profile of thorax not evenly rounded Pavement ant (Tetramorium “species E”) Pheidole californica Argentine Ant (Linepithema humile), subfamily Dolichoderinae Exotic species 3-4 mm in length Deep brown to light black Move rapidly in distinct trails Feed on honeydew Shallow nests (2 inches from soil surface) Alex Wild Does not bite or sting Carpenter Ant (Camponotus spp.), subfamily Formicinae Large ant: >6 mm in length Dark color with smooth, rounded thorax Workers most active at dusk and night One of most abundant and widespread genera worldwide Generalist scavengers and predators: feed on dead and living insects, nectar, fruit juices and Jack K.