Long-Term Field Evaluation of Mecinus Janthinus Releases Against
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

An Evaluation of Walleye in the Missouri River Between Holter Dam and Great Falls, Montana
An Evaluation of Walleye in the Missouri River between Holter Dam and Great Falls, Montana PPL-Montana MOTAC projects 771-09, 771-10, 759-11, 771-11 and Fisheries Bureau Federal Aid Job Progress Report Federal Aid Project Number F-113-R9, R10, R11, R12 Montana Statewide Fisheries Management Submitted to PPL-Montana 336 Rainbow Dam Great Falls, Mt. 59404 Prepared by Grant Grisak, Brad Tribby and Adam Strainer Montana Fish, Wildlife & Parks 4600 Giant Springs Road Great Falls, Mt. 49505 January 2012 1 Table of Contents Introduction…………………………………………………………………………… 5 Study Area……………………………………………………………………………. 5 Creel survey………………………………………………………………… 10 Angling……………………………………………………………………... 10 Fish Abundance………………………………………………………………………. 11 Tagging……………………………………………………………………………….. 15 Radio Telemetry……………………………………………………………………… 17 Early Life History…………………………………………………………………….. 28 Diet…………………………………………………………………………………… 32 Discussion…………………………………………………………………………….. 34 References……………………………………………………………………………. 37 2 List of Tables No. Page 1. Angler use statistics for Missouri River section 9, 1991-2009………………... 8 2. Economic statistics for the Missouri River section 9, 1995-2009……………... 9 3. Angler use statistics for Missouri River section 8, combined angler days with 9 section 9, and economic statistics for section 8 and section 9, 1991- 2009……………………………………………………………………………. 4. Landmarks and associated river miles in the Missouri River between Holter 18 Dam and Black Eagle Dam……………………………………………………. 5. Meristics of radio tagged walleye in Missouri River, total miles traveled and 26 total days radio transmitter was active, 2008-2011……………………………. 6. Locations in the Missouri River and proportional use by radio tagged walleye 27 2008-10. Missouri River, Montana……………………………………………. 7. Young of the year walleye seined at sites in the Missouri River between 30 Cascade and Great Falls……………………………………………………….. 8. Number of fish species sampled by year and total number of sites where 31 found. -

Little Muddy Creek Ranch 0 Old Ulm Cascade Road, Cascade, MT 59421
Little Muddy Creek Ranch 0 Old Ulm Cascade Road, Cascade, MT 59421 Offered at $3,750,000 Presented Exclusively by Trampus Corder, Broker Staci Corder, Broker 833-783-3224 toll free 406-622-3224 office [email protected] Property Information Acres: 3,647.77 Deeded Taxes: $4,350.00 (2020) Legal: Request from listing agent Water: Little Muddy Creek; Artesian well; 1 drilled well with solar pump; 4 developed springs; Additional springs on property that could be developed Utilities: Power on property Fencing: Perimeter and cross fenced; 6 separate pastures Outbuildings: Corral with ramp built in 2015 Scan the code for a virtual tour Access:Fort Benton Public Farm road access Chouteau County, Montana, 1280.0 AC +/- experience! Elevation: 3,370’ CCRs/HOAs: None Hunting District: 421; Several 80”+ antelope have been harvested on this land, including one that is top ten Pope and Young for Montana in bow hunting records! Executive Summary Among the squares of farm ground patched together constructing a quilt of the high productive, grain producing countryside. Little Muddy Creek flows through 3,647 alluring acres! The wonderous wetlands are located between Ulm and Cascade. The land is free of covenants and restrictions providing wide open space to roam, farm or ranch. With plenty of luscious grasses, the fairly flat land is ideal to open up grazing to your livestock. Watch the calves gain weight daily as they are enriched with the incomparable nutrient-dense native grasses that flourish along the lush riparian zones of North Central Montana. Unload and load cattle with the ramp on the corral. -

Missouri Riverside Lodge with Acreage
Missouri Riverside Lodge With Acreage 3103 Old US Highway 91, Cascade, MT $850,000 Photography courtesy of Leonard (Arnie) Gidlow© Presented Exclusively by Trampus Corder, Broker Staci Corder, Broker 406-622-3224 office [email protected] 1312 Front Street, Fort Benton, MT CorderandAssociates.com Property Information Acres: 23.97 Taxes: $6,544.33 (2019) Personal Property: Complete inventory list upon request Legal: S20, T16 N, R02 W, IN SENE S/RD, TR 2 COS 4948 MK 18; S20, T16 N, R02 W, 3870, PARCEL 000, TR B, COS #3870, IN SW4NE4 MK 28B Executive Summary The twinkling Montana stars have aligned to create a flawless opportunity to acquire a legendary lodge located on the most desirable section of the Missouri River that is world renowned for its exceptional trout fishing and infinite outdoor recreating possibilities. This spectacular lodge consisting of six guest rooms is quiet and peaceful giving visitors an authentic and tranquil Montana escape. The lodge offers just under 24 acres of the vast mountainous terrain to roam and experience the natural wonders that so many have fallen in love with. The crisp,Fort Benton clear Farmwaters of the Missouri River are calm and serene setting the tone for a relaxing float until adrenaline burstsChouteau throughout County, Montana, your 1280.0 body AC +/-when your fishing pole starts tugging implying something large at the other end. Photographs become the sacred proof of the several fish that are netted 10 inches in length or more in a section of river that boasts 7000+ fish per mile. The cold weather will not allow any disappointment to you or your guests as fishing year-round transpires with the same awesome rewards. -

Montana Historical Society Preserves Montana’S Treasures
Montana Historical Society preserves Montana’s treasures Historic Preservation Working to save historic sites, including the Doncaster Round Barn nearTwin Bridges Museum Exhibits like the new C. M. Russell gallery— showcasing the work of Montana’s favorite artist Education and Outreach Educational resources for children and adults, such as Archeology Day and the new Montana history textbook Publications Montana The Magazine of Western History, published since 1951, and books by the Montana Historical Society Press Research Center Serving the public with professional assistance and vast Montana resource materials Mike Cooney Denise C. King Interim Director Centralized Services 444-5485 Division Administrator 225 North Roberts, Helena, MT 444-4699 (406) 444-2694 montanahistoricalsociety.org Services Provided to the Citizens of the Treasure State by the Montana Historical Society, 2007 to 2010 (Data arranged by county) 2 Montana Historical Society Statewide Services The five operational programs of the Montana Historical Society— Education and Outreach, Historic Preservation, Museum, Research Center, and Publications—provide essential services to Treasure State citizens living in all of Montana’s fifty-six counties. Listed below is a brief summary of just a few of the Society’s statewide offerings between 2007 and 2010. On the pages that follow, more detail is given in regard to some of the specific services rendered to the residents of each county. • The Big Read: This program, funded through grants from the National Endowment for the Arts and the Institute of Museum and Library Services, was designed to reach rural middle and high school students. It served seventeen schools across the state, providing books and bringing public programs to these communities and MHS services directly to the classrooms. -

Missouri Riverside Lodge
Missouri Riverside Lodge 3103 Old US Highway 91, Cascade, MT $675,000 Photography courtesy of Leonard (Arnie) Gidlow© Presented Exclusively by Trampus Corder, Broker Staci Corder, Broker 406-622-3224 office [email protected] 1312 Front Street, Fort Benton, MT CorderandAssociates.com Property Information Acres: 13.96 Taxes: $6,169.49 (2019) Personal Property: Complete inventory list upon request Legal: S20, T16 N, R02 W, IN SENE S/RD, TR 2 COS 4948 MK 18 Executive Summary The twinkling Montana stars have aligned to create a flawless opportunity to acquire a legendary lodge located on the most desirable section of the Missouri River that is world renowned for its exceptional trout fishing and infinite outdoor recreating possibilities. This spectacular lodge consisting of six guest rooms is quiet and peaceful giving visitors an authentic and tranquil Montana escape. The main area of the lodge consists of five components: shop/office, main lodge area, mechanical area, upstairs apartment, and patio. The lodge sits on 13.96 acres with views of the beautiful mountainous terrain. The crisp, clear waters of the Missouri River are calm and serene setting the tone for a relaxingFort Benton float Farm until adrenaline bursts throughout your body when your fishing pole starts tugging implying somethingChouteau County, large Montana, at the 1280.0 other AC end.+/- Photographs become the sacred proof of the several fish that are netted 10 inches in length or more in a section of river that boasts 7000+ fish per mile. The cold weather will not allow any disappointment to you or your guests as fishing year-round transpires with the same awesome rewards. -
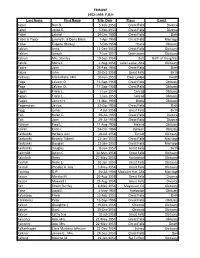
1952-1959 F-G-H Last Name First Name Trib Date Place Event Fabel Don D
TRIBUNE 1952-1959 F-G-H Last Name First Name Trib Date Place Event Fabel Don D. 5-Feb-1953 Great Falls Divorce Fabel Joyce E. 5-Feb-1953 Great Falls Divorce Fabel Leland 24-Oct-1955 Great Falls Birth Fabel & Yoder Leland F. & Doris Marie 1-Apr-1955 Great Falls Marriage Faber Eugene Stanley 7-Oct-1956 Havre Obituary Fabian Anna 11-Dec-1952 Great Falls Obituary Fabian Joseph 7-Jun-1957 Lewistown Obituary Fabian Mrs. Stanley 19-Sep-1954 Belt Birth of Daughter Fabila Maria L 1-Aug-1954 Lake Louise, Alta Obituary Fabre John 25-Feb-1956 Great Falls Birth Fabre John 20-Oct-1958 Great Falls Birth Fadness Clara Marie, Mrs. 16-Jun-1953 Deer Lodge Death Faga LaVern O. 16-Sep-1955 Great Falls Obituary Faga LaVern O. 17-Sep-1955 Great Falls Obituary Fagan Frank L. 1-Jun-1954 Conrad Obituary Fagan Frank L. 1-Jun-1953 Conrad Obituary Fagan James H. 11-Mar-1956 Brady Obituary Fagenstrom James 22-Oct-1956 Great Falls Birth Fagg James 4-Jul-1958 Great Falls Birth Fah Helen C. 29-Jul-1955 Great Falls Divorce Fah John 29-Jul-1955 Great Falls Divorce Fahrion Fred L. 11-Aug-1956 Helena Obituary Fairall Earl H. 24-Oct-1956 Opheim Obituary Fairbanks Barbara Ann 10-Jul-1954 Turner Obituary Fairbanks Beverly Tidwell 22-Jan-1953 Great Falls Marriage Fairbanks Douglas 22-Jan-1953 Great Falls Marriage Fairbanks Douglas 8-Jun-1957 Great Falls Birth Fairchild Byron C. 30-Mar-1958 Great Falls Obituary Faircloth Harry 27-May-1954 Harlowton Obituary Fairfull Nellie L. -

Minutes of the Meeting Taxation Committee Montana State Senate
MINUTES OF THE MEETING TAXATION COMMITTEE MONTANA STATE SENATE April 1, 1987 The fifty-fifth meeting of the Senate Taxation Committee was called to order at 8:00 A.M. on April 1, 1987 by Chairman George McCallum in Room 325 of the Capitol Building. ROLL CALL: All committee members were present. CONSIDERATION OF HB 84: Representative Winslow, House District 89, presented this bill to the committee. We are putting $1.2 million a year into a tourism promotion program when the states around us are putting $5 million. We are 49th in the nation as far as attraction and tourism. This bill has been wo~ked on for a year and some changes were made and some commitments were made from people that previously have not supported this kind of a tax. Several people have looked at ~hat could be done to put Montana in a position of c~mpetition in tourism. If we do not compete we lose. Montana does not even have an opportunity to get the word out on what a wonderful place Montana would be to vacation, people come through Montana because that is where the road goes, not because they have heard about it. This bill assesses a 4% bed tax on motels and campgrounds. The House committee added a couple of amendments which he supports. One would have a percentage go to the Historical Society, which plays a critical roll. The other breaks some of it down to go to the university system. John Wilson from the Department has some clean up amendments for some of the action taken in the House. -

United States Department of the Interior National Park Service NATIONAL REGISTER of HISTORIC PLACES CONTINUATION SHEET Section
NPS Form 10-900a OMB No. 1024-0018 (8-86) United States Department of the Interior National Park Service NATIONAL REGISTER OF HISTORIC PLACES CONTINUATION SHEET Section ___ Page __ SUPPLEMENTARY LISTING RECORD NRIS Reference Number: 09001180 Date Listed: 1/4/2010 Hardy Bridge Cascade MT Property Name County State Montana 7 s Historic Steel Truss Bridges MPS Multiple Name This property is listed in the National Register of Historic Places in accordance with the attached nomination documentation subject to the following exceptions, exclusions, or amendments, notwithstanding the National Park Service certification included in the nomination documentation. lature/of the Keeper Date of Action Amended Items in Nomination: Classification: The Number of Contributing Properties Previously Listed should read: 0 [This refers only to resources within the nominated boundaries of this property not to other bridge locations associated with the MPS context.] These clarifications were confirmed with the MT SHPO office. DISTRIBUTION: National Register property file Nominating Authority (without nomination attachment) NPS Form 10-900 (Rev. 01/2009) OMBNo. 1024-0018 United States Department of the Interior National Park Service NOV 202009 National Register of Historic Places Registration Form This form is for use in nominating or requesting determinations for individual properties and districts. See instructions in National Register Bulletin, How to Complete the National Register of Historic Places Registration Form. If any item does not apply to the property being documented, enter "N/A" for "not applicable." For functions, architectural classification, materials, and areas of significance, enter only categories and subcategories from the instructions. Place additional certification comments, entries, and narrative items on continuation sheets (NPS Form 10-900a). -

Fourth Biennial Report to the State Preservation Review Board by the Montana Department of Transportation As Specified by Senate Bill No
FOURTH BIENNIAL REPORT TO THE STATE PRESERVATION REVIEW BOARD BY THE MONTANA DEPARTMENT OF TRANSPORTATION AS SPECIFIED BY SENATE BILL NO. 3 (2011) ON THE STATUS AND NEEDS OF AGENCY HERITAGE PROPERTIES January 2018 Report Prepared By: Jon Axline Environmental Services Montana Department of Transportation 2701 Prospect Avenue Helena, Montana 59620-1001 FOURTH BIENNIAL REPORT TO THE STATE PRESERVATION REVIEW BOARD BY THE MONTANA DEPARTMENT OF TRANSPORTATION AS SPECIFIED BY SENATE BILL NO. 3 (2011) ON THE STATUS AND NEEDS OF AGENCY HERITAGE PROPERTIES January 2018 Introduction This document constitutes the Montana Department of Transportation’s (MDT) fourth biennial report to the Historic Preservation Review Board as required under the 2011 Montana Senate Bill No. 3 (the State Heritage Stewardship amendment to the Montana Antiquities Act), a regulation that requires State Agencies and the Montana University System to biennially report to the Board on the status and maintenance needs of agency heritage properties. This report includes the MDT’s known heritage properties, their status, condition, and priority for preservation. In 2016, the MDT reported 108 heritage properties owned by the agency. Since that reporting period, nine properties have been removed from MDT ownership or demolished and thirteen properties added to the list. The new properties consist primarily of MDT-owned airway beacons. In 2018, the MDT’s heritage properties list includes 119 historic properties. Most of the heritage properties detailed in this report were documented and treated under Section 106 (36CFR 800) of the National Historic Preservation Act (1966) as part of the MDT’s cultural resource program. That regulation is applicable to federally-funded projects that have the potential to impact historic properties. -
Macroinvertebrate Monitoring for the Upper Missouri River: Building a Long-Term Data Set 2015-2019
UMOWA Missouri River Project Baseline Aquatic Surveys Montana Biological Survey April 2020 Macroinvertebrate Monitoring for the Upper Missouri River: Building a Long-term Data Set 2015-2019 Prepared for: Alan Shaw, Sherry Meador, Pete Petersen, Pat Hunter and Bailey Sory Project Managers/Co-Chairs Missouri River Monitoring Station below the Dearborn River Fall 2019 Prepared by: David Stagliano, Aquatic Ecologist Montana Biological Survey Helena, Montana April 2020 UMOWA Missouri River Project Baseline Aquatic Macroinvertebrate Surveys Montana Biological Survey April 2020 Table of Contents Acknowledgements .................................................................................................................................... 3 Executive Summary ................................................................................................................................... 4 1.0 Introduction ........................................................................................................................................... 6 2.0 Methods ................................................................................................................................................ 8 2.1 Habitat and Physical Water Sampling .......................................................................................... 8 2.2 Macroinvertebrate Sampling .......................................................................................................... 8 2.3 Taxonomic Analysis ....................................................................................................................... -
Bulletin Summer 2017
VOLUME 46 NEWSLETTER SUMMER 2017 Downtown Bozeman 1875 GALLATIN MUSEUM photo Montana Ghost Town The Prez Sez TERRY HALDEN Quarterly The Montana Ghost Town Quarterly is published four times a year by the Convention 2017 in Bozeman is all set to go and it looks like Montana Ghost Town Preservation Society, it will be another winner. Margie Kankrik and Marilyn Murdock have P.O. Box 1861, Bozeman, Montana 59771. e-mail: [email protected] done a tremendous job in putting it together, along with some exciting www.mtghosttown.org guest speakers that are sure to inform and entertain you. Next year it Copyright © 2017, all rights reserved. is Darian Halden’s turn to put a convention together. I can assure Founded in 1970, the Montana Ghost Town you, from past experience, it is not an easy task. Preservation Society is a 501c3 non-profit organization dedicated to educating the public to In this issue of the newsletter, we welcome back Rachel the benefits of preserving the historic buildings, Phillips who has written an article about the Bozeman Hot Springs, sites, and artifacts that make up the living history of Montana. which is not as well-known as those hot springs between Butte and Opinions expressed in the bylined articles are Anaconda, but nevertheless, sports just as interesting a history. Your the authors’ and do not necessarily represent Vice-President, Brad O’Grosky has written an article about the history the views of the M. G. T. P. S. of bridges in Montana. Although they do not have the romance of SUMMER 2017 ghost towns, they were just as important to the development of Montana and as such their history should be maintained. -
Hardy Creek Hideaway 47 Prewett Creek, Cascade, MT, 59421
Hardy Creek Hideaway 47 Prewett Creek, Cascade, MT, 59421 Offered at $325,000 Presented Exclusively by Mark Daly, REALTOR® 406-622-3224 office 406-459-9642 cell [email protected] Property Information Acres: 184 Deeded Taxes: $376.00 (2020) Legal: PANORAMA MTNS ISLAND VIEW, S17, T24N, R31W, Lot 15, ACRES 1.53 Utilities: None; Power is very far away; Generators, wind or hydro would be acceptable means Covenants/HOAs: None Water: Hardy Creek traverses property; Unrecorded well Water rights: Yes Outbuildings/Structures: 3 older shells of cabins Hunting District: 421; Hunting for bears, elk, deer, mountainFort Benton lions Farm Chouteau County, Montana, 1280.0 AC +/- Executive Summary Lush, verdant grass encases the gently rolling hills of this 184 acres in the Dearborn Valley. Through the center, a blue ribbon of Hardy Creek flows toward the sky- punching mountains in the background. Wild foods grow abundantly for grazing wildlife as the climate is mild and dreamy. The endless, jewel-blue sky is a feast for the eyes. Escape the sounds of honking horns and enjoy the tranquil sounds of plinking water over the rocky bottom of Hardy Creek. The mountain air ignites the skin with perky goosebumps. Some of the best of Montana hunting and fishing happens in this area. The southern chunk of this property borders state land increasing the opportunity to seek out adventure. As cheery swallows squeak, wildflowers radiate a sweet scent as they are tickled by the whispering winds. The Aspens, Willows and Spruce provide shelter for animal life and the waters of the valley are worthy of your time offering trophy catches from the blue ribbon rivers.