Community Structure, Life Histories and Secondary Production of Stoneflies in Two Small Mountain Streams with Different Degree of Forest Cover
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Monte L. Bean Life Science Museum Brigham Young University Provo, Utah 84602 PBRIA a Newsletter for Plecopterologists
No. 10 1990/1991 Monte L. Bean Life Science Museum Brigham Young University Provo, Utah 84602 PBRIA A Newsletter for Plecopterologists EDITORS: Richard W, Baumann Monte L. Bean Life Science Museum Brigham Young University Provo, Utah 84602 Peter Zwick Limnologische Flußstation Max-Planck-Institut für Limnologie, Postfach 260, D-6407, Schlitz, West Germany EDITORIAL ASSISTANT: Bonnie Snow REPORT 3rd N orth A merican Stonefly S ymposium Boris Kondratieff hosted an enthusiastic group of plecopterologists in Fort Collins, Colorado during May 17-19, 1991. More than 30 papers and posters were presented and much fruitful discussion occurred. An enjoyable field trip to the Colorado Rockies took place on Sunday, May 19th, and the weather was excellent. Boris was such a good host that it was difficult to leave, but many participants traveled to Santa Fe, New Mexico to attend the annual meetings of the North American Benthological Society. Bill Stark gave us a way to remember this meeting by producing a T-shirt with a unique “Spirit Fly” design. ANNOUNCEMENT 11th International Stonefly Symposium Stan Szczytko has planned and organized an excellent symposium that will be held at the Tree Haven Biological Station, University of Wisconsin in Tomahawk, Wisconsin, USA. The registration cost of $300 includes lodging, meals, field trip and a T- Shirt. This is a real bargain so hopefully many colleagues and friends will come and participate in the symposium August 17-20, 1992. Stan has promised good weather and good friends even though he will not guarantee that stonefly adults will be collected during the field trip. Printed August 1992 1 OBITUARIES RODNEY L. -

Download .PDF(1340
Stark, Bill P. and Stephen Green. 2011. Eggs of western Nearctic Acroneuriinae (Plecoptera: Perlidae). Illiesia, 7(17):157-166. Available online: http://www2.pms-lj.si/illiesia/Illiesia07-17.pdf EGGS OF WESTERN NEARCTIC ACRONEURIINAE (PLECOPTERA: PERLIDAE) Bill P. Stark1 and Stephen Green2 1,2 Box 4045, Department of Biology, Mississippi College, Clinton, Mississippi, U.S.A. 39058 1 E-mail: [email protected] 2 E-mail: [email protected] ABSTRACT Eggs for western Nearctic acroneuriine species of Calineuria Ricker, Doroneuria Needham & Claassen and Hesperoperla Banks are examined and redescribed based on scanning electron microscopy images taken from specimens collected from a substantial portion of each species range. Within genera, species differences in egg morphology are small and not always useful for species recognition, however eggs from one population of Calineuria are significantly different from those found in other populations and this population is given informal recognition as a possible new species. Keywords: Plecoptera, Calineuria, Doroneuria, Hesperoperla, Egg morphology, Western Nearctic INTRODUCTION occur in the region (Baumann & Olson 1984; Scanning electron microscopy (SEM) is often used Kondratieff & Baumann 2002; Stark 1989; Stark & to elucidate chorionic features for stoneflies (e.g. Gaufin 1976; Stark & Kondratieff 2004; Zuellig et al. Baumann 1973; Grubbs 2005; Isobe 1988; Kondratieff 2006). SEM images for eggs of the primary western 2004; Kondratieff & Kirchner 1996; Nelson 2000; acroneuriine genera, Calineuria Ricker, Doroneuria Sivec & Stark 2002; 2008; Stark & Nelson 1994; Stark Needham & Claassen and Hesperoperla Banks include & Szczytko 1982; 1988; Szczytko & Stewart 1979) and single images for each of these genera in Stark & Nearctic Perlidae were among the earliest stoneflies Gaufin (1976), three images of Hesperoperla hoguei to be studied with this technique (Stark & Gaufin Baumann & Stark (1980) and three images of H. -

Life Cycle of Three Stonefly Species (Plecoptera) from an Apenninic Stream (Italy) with the Description of the Nymph of Nemoura Hesperiae
Life cycle of three stonefly species (Plecoptera) from an Apenninic stream (Italy) with the description of the nymph of Nemoura hesperiae Tierno de Figueroa J.M. (1), Bo T. (2), López-Rodríguez M.J. (1), Fenoglio S. (2) (1) Universidad de Granada, Departamento de Biología Animal, 18071, Granada, Spain. Email: [email protected], [email protected]. (2) Università del Piemonte Orientale, Dipartimento di Scienze dell’Ambiente e della Vita, Via Bellini 25, 15100, Alessandria, Italy. Email: [email protected], [email protected]. Running title: Life cycle of Plecoptera from an Apenninic stream 1 Abstract The life cycles of Isoperla grammatica, Amphinemura sulcicollis and Nemoura hesperiae are studied in a Northern Italy stream, sited in the Apennines. The three species show a univoltine life cycle and their development coincides approximately in the same period of the year, although the one of I. grammatica is longer. Possible egg dormancy could exist in the A. sulcicollis and N. hesperiae life cycles, but this needs to be proved. The three taxa show a spring flight period in the study area. Growth is almost constant through the life cycle, except in A. sulcicollis in which an increase is observed at the end of the development. N. hesperiae exhibits a faster growth than the other species. In addition, in this paper, the nymph of N. hesperiae is described and designed for the first time. Résumé Nous avons étudiés les cycles de vie de Isoperla grammatica, Amphinemura sulcicollis et Nemoura hesperiae dans une riviere des Apennins, dans l’Italie septentrional. Les trois espèces ont des cycles univoltins synchronisés a peu prés dans le même période de l’année, même si le cycle de I. -
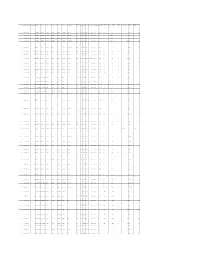
Appendix D: Summary of Accepted ECOTOX Papers
Effect Dur Dur Unit Dur Dur Unit Conc Conc Units Conc Value1 Purity CAS Number Chemical Name Species Number Phylum Class Order Family Genus Species Common Name Group Effect Meas Endpt1 Endpt2 Habitat Plant/Animal Media Orig Orig Preferred Preferred Type Conc Value1 Orig Orig Adjusted 1 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BEH AVO STIM NOAEL terrestrial Animal NONE 30 d 30 d F 2 mg/kg/d 1.96 2 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM LIPD LOAEL aquatic Animal FW 1 h 4.17E-02 d A 4 mg/L 4 3 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM PRCO LOAEL aquatic Animal FW 1 h 4.17E-02 d A 4 mg/L 4 4 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM LIPD LOAEL aquatic Animal FW 1 h 4.17E-02 d A 10.5 mg/L 10.5 5 298044 Disulfoton 4913 Chordata Mammalia Rodentia Muridae Mus musculus House mouse BCM BCM HXBT LOAEL terrestrial Animal NONE 3 d 3 d F 35.1 uM/kg 35.1 6 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM LOAEL terrestrial Animal NONE 24 h 1 d F 0.26 mg/kg 0.234 7 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM IC50 terrestrial Animal NONE 1 wk 7 d F 0.6 ppm 0.6 8 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM NOAEL LOAEL terrestrial Animal NONE -

In Baltic Amber 85
Overview and descriptions of fossil stoneflies (Plecoptera) in Baltic amber 85 Entomologie heute 22 (2010): 85-97 Overview and Descriptions of Fossil Stoneflies (Plecoptera) in Baltic Amber Übersicht und Beschreibungen von fossilen Steinfliegen (Plecoptera) im Baltischen Bernstein CELESTINE CARUSO & WILFRIED WICHARD Summary: Three new fossil species of stoneflies (Plecoptera: Nemouridae and Leuctridae) from Eocene Baltic amber are being described: Zealeuctra cornuta n. sp., Lednia zilli n. sp., and Podmosta attenuata n. sp.. Extant species of these three genera are found in Eastern Asia and in the Nearctic region. It is very probably that the genera must have been widely spread across the northern hemisphere in the Cretaceous period, before Europe was an archipelago in Eocene. The current state of knowledge about the seventeen Plecoptera species of Baltic amber is shortly presented. Due to discovered homonymies, the following nomenclatural corrections are proposed: Leuctra fusca Pictet, 1856 in Leuctra electrofusca Caruso & Wichard, 2010 and Nemoura affinis Berendt, 1856 in Nemoura electroaffinis Caruso & Wichard, 2010. Keywords: Fossil insects, fossil Plecoptera, Eocene, paleobiogeography Zusammenfassung: In dieser Arbeit werden drei neue fossile Steinfliegen-Arten (Plecoptera: Nemouridae und Leuctridae) des Baltischen Bernsteins beschrieben: Zealeuctra cornuta n. sp., Lednia zilli n. sp., Podmosta attenuata n. sp.. Rezente Arten der drei Gattungen sind in Ostasien und in der nearktischen Region nachgewiesen. Sehr wahrscheinlich breiteten sich die Gattungen in der Krei- dezeit über die nördliche Hemisphäre aus, noch bevor Europa im Eozän ein Archipel war. Der gegenwärtige Kenntnisstand über die siebzehn Plecoptera Arten des Baltischen Bernsteins wird kurz dargelegt. Wegen bestehender Homonymien werden folgende nomenklatorische Korrekturen vorgenommen: Leuctra fusca Pictet, 1856 in Leuctra electrofusca Caruso & Wichard, 2010 und Nemoura affinis Berendt, 1856 in Nemoura electroaffinis Caruso & Wichard, 2010. -

Plecoptera: Perlidae), with an Annotated Checklist of the Subfamily in the Realm
Opusc. Zool. Budapest, 2016, 47(2): 173–196 On the identity of some Oriental Acroneuriinae taxa (Plecoptera: Perlidae), with an annotated checklist of the subfamily in the realm D. MURÁNYI1 & W.H. LI2 1Dávid Murányi, Department of Civil and Environmental Engineering, Ehime University, Bunkyo-cho 3, Matsuyama, 790-8577 Japan, and Department of Zoology, Hungarian Natural History Museum, H-1088 Budapest, Baross u. 13, Hungary. E-mail: [email protected], [email protected] 2Weihai Li, Department of Plant Protection, Henan Institute of Science and Technology, Xinxiang, Henan, 453003 China. E-mail: [email protected] Abstract. The monotypic Taiwanese genus Mesoperla Klapálek, 1913 is redescribed on the basis of a male syntype specimen, and its affinities are re-evaluated. The single female type specimen of further two Oriental monotypic genera, Kalidasia Klapálek, 1914 and Nirvania Klapálek, 1914, are confirmed to be lost or destroyed respectively; both genera are considered as nomina dubia. The Sichuan endemic Acroneuria grahami Wu & Claassen, 1934 is redescribed on the basis of male holotype. Distinctive characters of the genus Brahmana Klapálek, 1914 consisting of five, inadequately known Oriental species are discussed. Flavoperla needhami (Klapálek, 1916) and Sinacroneuria sinica (Yang & Yang, 1998) comb. novae are suggested for an Indian species originally described in Gibosia Okamoto, 1912 and a Chinese species originally described in Acroneuria Pictet, 1841. At present, 62 species of Acroneuriinae, classified in 10 valid genera are reported from the Oriental Realm but 29 species are inadequately known. A key is presented to distinguish males of the Asian Acroneuriinae genera. Asian distribution of each genera are detailed and depicted on a map. -

Plecoptera: Nemouridae), New Stonefly Species from the Eastern Nearctic Region and Notes on the Life Cycle of S
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Illiesia Jahr/Year: 2006 Band/Volume: 02 Autor(en)/Author(s): Grubbs Scott A. Artikel/Article: Soyedina alexandria and S. calcarea (Plecoptera: Nemouridae), new stonefly species from the Eastern Nearctic region and notes on the life cycle of S. calcarea. 39-49 Grubbs, Scott A. 2006. Soyedina alexandria and S. calcarea (Plecoptera: Nemouridae), new stonefly species from the eastern Nearctic region and notes on the life cycle of S. calcarea. Illiesia, 2(6):39‐49. Available online: http://www2.pms‐lj.si/illiesia/Illiesia02‐06.pdf SOYEDINA ALEXANDRIA AND S. CALCAREA (PLECOPTERA: NEMOURIDAE), NEW STONEFLY SPECIES FROM THE EASTERN NEARCTIC REGION AND NOTES ON THE LIFE CYCLE OF S. CALCAREA. Scott A. Grubbs Department of Biology and Center for Biodiversity Studies, Western Kentucky University, 1906 College Heights Boulevard 11080, Bowling Green, KY 42101‐1080, U.S.A. E‐mail: [email protected] ABSTRACT Two new species of eastern Nearctic Nemouridae, Soyedina alexandria and S. calcarea, are described from the karst regions of Tennessee and Kentucky, U.S.A. Illustrations and scanning electron micrographs of the male and female stages are provided and diagnostic characters separating these two species from closely‐related S. vallicularia are presented. Soyedina vallicularia was studied across its broad range to address potential morphological variation, which may impede the ability to differentiate S. alexandria and S. calcarea from S. vallicularia. A diagnostic key is provided for the males of S. vallicularia, S. alexandria, and S. calcarea. Nymphs of S. calcarea were collected monthly over a two‐year period from the Kentucky type locality and size frequency histograms of head capsule width revealed a univoltine‐slow cycle. -

Nanonemoura, a New Stonefly Genus from the Columbia River Gorge, Oregon (Plecoptera: Nemouridae)
Western North American Naturalist Volume 61 Number 4 Article 3 11-15-2001 Nanonemoura, a new stonefly genus from the Columbia River Gorge, Oregon (Plecoptera: Nemouridae) R. W. Baumann Brigham Young University G.R. Fiala Gresham, Oregon Follow this and additional works at: https://scholarsarchive.byu.edu/wnan Recommended Citation Baumann, R. W. and Fiala, G.R. (2001) "Nanonemoura, a new stonefly genus from the Columbia River Gorge, Oregon (Plecoptera: Nemouridae)," Western North American Naturalist: Vol. 61 : No. 4 , Article 3. Available at: https://scholarsarchive.byu.edu/wnan/vol61/iss4/3 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Western North American Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Western North American Naturalist 61(4), © 2001, pp. 403–408 NANONEMOURA, A NEW STONEFLY GENUS FROM THE COLUMBIA RIVER GORGE, OREGON (PLECOPTERA: NEMOURIDAE) R.W. Baumann1 and G.R. Fiala2 ABSTRACT.—Nanonemoura, a new genus of Nearctic Nemouridae, is described to accommodate Nemoura wahkeena Jewett. New descriptions and illustrations are provided for the adult male, adult female, and nymph collected at Wah- keena Spring, Columbia River George, Oregon. A diagnosis is furnished to distinguish the new genus from related gen- era of the subfamily Nemourinae. Key words: Plecoptera, Nemouridae, Nanonemoura, Columbia River Gorge, Oregon. Nearly 50 years ago Jewett (1954) named a nymph of this species. We discovered that the nemourid species from the Columbia River species does not live in the falls or even in the Gorge. -

Aquatic Insect Ecophysiological Traits Reveal Phylogenetically Based Differences in Dissolved Cadmium Susceptibility
Aquatic insect ecophysiological traits reveal phylogenetically based differences in dissolved cadmium susceptibility David B. Buchwalter*†, Daniel J. Cain‡, Caitrin A. Martin*, Lingtian Xie*, Samuel N. Luoma‡, and Theodore Garland, Jr.§ *Department of Environmental and Molecular Toxicology, Campus Box 7633, North Carolina State University, Raleigh, NC 27604; ‡Water Resources Division, U.S. Geological Survey, 345 Middlefield Road, MS 465, Menlo Park, CA 94025; and §Department of Biology, University of California, Riverside, CA 92521 Edited by George N. Somero, Stanford University, Pacific Grove, CA, and approved April 28, 2008 (received for review February 20, 2008) We used a phylogenetically based comparative approach to evaluate ecosystems today (e.g., trace metals) (6). This variation in the potential for physiological studies to reveal patterns of diversity susceptibility has practical implications, because the ecological in traits related to susceptibility to an environmental stressor, the structure of aquatic insect communities is often used to indicate trace metal cadmium (Cd). Physiological traits related to Cd bioaccu- the ecological conditions in freshwater systems (7–9). Differ- mulation, compartmentalization, and ultimately susceptibility were ences among species’ responses to environmental stressors can measured in 21 aquatic insect species representing the orders be profound, but it is uncertain whether the cause is related to Ephemeroptera, Plecoptera, and Trichoptera. We mapped these ex- functional ecology [usually the assumption (10, 11)] or physio- perimentally derived physiological traits onto a phylogeny and quan- logical traits (5, 12–14), which have received considerably less tified the tendency for related species to be similar (phylogenetic attention. To the degree that either is involved, their link to signal). -

Nymphs of North American Perlodinae Genera (Plecoptera: Perlodidae)
Great Basin Naturalist Volume 44 Number 3 Article 1 7-31-1984 Nymphs of North American Perlodinae genera (Plecoptera: Perlodidae) Kenneth W. Stewart North Texas State University, Denton, Texas Bill P. Stark Mississippi College, Clinton, Mississippi Follow this and additional works at: https://scholarsarchive.byu.edu/gbn Recommended Citation Stewart, Kenneth W. and Stark, Bill P. (1984) "Nymphs of North American Perlodinae genera (Plecoptera: Perlodidae)," Great Basin Naturalist: Vol. 44 : No. 3 , Article 1. Available at: https://scholarsarchive.byu.edu/gbn/vol44/iss3/1 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Great Basin Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. The Great Basin Naturalist Published at Provo, Utah, by Brigham Young University ISSN 0017-3614 Volume 44 July 31, 1984 No. 3 NYMPHS OF NORTH AMERICAN PERLODINAE GENERA (PLECOPTERA: PERLODIDAE)' Kenneth VV. Stewart- and Bill P. Stark' Abstract.— Nymphs of the type or other representative species of the 22 North American Perlodinae genera are comparatively described and illustrated for the first time. The first complete generic key for the subfamily incorporates recent nymph discoveries and revisions in classification. References to all previous nymph descriptions and illustrations and major life cycle and food habits studies are given for the 53 North American species in the subfamilv, and a listing of species and their current distributions by states and provinces is provided for each genus. The previously unknown nymph of Chcrnokrihts misnomus is described and illustrated. -

Phenology and Diversity of Adult Stoneflies (Plecoptera) of a Small Coastal Stream, California
Bottorff, Richard L., Loren D. Bottorff, 2007. Phenology and diversity of adult stoneflies (Plecoptera) of a small coastal stream, California. Illiesia, 3(1):1‐9. Available online: http://www2.pms‐lj.si/illiesia/Illiesia03‐01.pdf PHENOLOGY AND DIVERSITY OF ADULT STONEFLIES (PLECOPTERA) OF A SMALL COASTAL STREAM, CALIFORNIA Richard L. Bottorff1 and Loren D. Bottorff 2 11963 Toppewetah Street, South Lake Tahoe, CA 96150, E‐mail: [email protected] 23265 Sandhurst Court, Cameron Park, CA 95682, E‐mail: [email protected] ABSTRACT Collections of adult stoneflies over a full year at Irish Gulch Creek, Mendocino Co., California, revealed 23 species. Adults were present at all times of the year. Species number varied from an autumnal low of 2 to a spring peak of 13. Adults of most species were present for less than 3 months, but Malenka depressa adults were present year‐round. Hesperoperla hoguei was the only strictly autumnal‐emerging species. The report of Suwallia dubia from Irish Gulch Creek represents a new California record. The stonefly faunas of Irish Gulch Creek (low coastal) and Sagehen Creek (high Sierra Nevada) were compared. Both creeks had similar numbers of species, but the species composition differed greatly, reflecting dissimilar environments (elevation, water temperature, thermal accumulation, and discharge). Irish Gulch Creek had uniform warmer temperatures; Sagehen Creek had variable colder temperatures. Peak emergence at Irish Gulch Creek occurred 2 months earlier than at Sagehen Creek. Keywords: Plecoptera, seasonal flight period, biodiversity, thermal stability, North Coast bioregion INTRODUCTION we studied these aspects for a small stream on the As might be expected from its varied topography, north coast of California. -

F- Oliver Gathmann a Thesis Submitted in Conformity with the Requirements
INSECT COMMUNITY PATTERNS M SPACE: STATISTICAL TOOLS, SIMULATIONS, AND EMPIRICAL DATA FROM SOUTHERNONTARIO COLDWATER SPIUNGS F-Oliver Gathmann A thesis submitted in conformity with the requirements for the degree of Ph.D. Graduate Department of Zoology University of Toronto Copyright @ 2000 by F. Oliver Gathmann National Library Bibliothèque nationale du Canada Acquisitions and Acquisitions et Bibliographie Services services bibliographiques 395 Wellington Street 395, rue Wellington OttawaON K1AON4 Ottawa ON K1A ON4 Canada Canada The author has granted a non- L'auteur a accorde une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or sell reproduire, prêter, distribuer ou copies of this thesis in microform, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/nlm, de reproduction sur papier ou sur format électronique. The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts from it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation. Contents Prolog 1 Bibliography .................................................. 6 1 Empirical Data 1 Insect emergence in Southern Ontario coldwater springs: patterns of species diversity and species- environment relationships 9 1.1 introduction ................................................ 9 1.2 StudyArea ................................................ 11 1.3 Material and Methods ........................................... 12 1.4 Results ................................................... 13 1.4.1 Abioticdata ............................................ 13 1.4.2 Taxonlist ............................................. 18 1.4.3 Species frequency .......................................