Some Evolutionary Trends in Plecoptera
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Aquatic Insects Are Dramatically Underrepresented in Genomic Research
insects Communication Aquatic Insects Are Dramatically Underrepresented in Genomic Research Scott Hotaling 1,* , Joanna L. Kelley 1 and Paul B. Frandsen 2,3,* 1 School of Biological Sciences, Washington State University, Pullman, WA 99164, USA; [email protected] 2 Department of Plant and Wildlife Sciences, Brigham Young University, Provo, UT 84062, USA 3 Data Science Lab, Smithsonian Institution, Washington, DC 20002, USA * Correspondence: [email protected] (S.H.); [email protected] (P.B.F.); Tel.: +1-(828)-507-9950 (S.H.); +1-(801)-422-2283 (P.B.F.) Received: 20 August 2020; Accepted: 3 September 2020; Published: 5 September 2020 Simple Summary: The genome is the basic evolutionary unit underpinning life on Earth. Knowing its sequence, including the many thousands of genes coding for proteins in an organism, empowers scientific discovery for both the focal organism and related species. Aquatic insects represent 10% of all insect diversity, can be found on every continent except Antarctica, and are key components of freshwater ecosystems. However, aquatic insect genome biology lags dramatically behind that of terrestrial insects. If genomic effort was spread evenly, one aquatic insect genome would be sequenced for every ~9 terrestrial insect genomes. Instead, ~24 terrestrial insect genomes have been sequenced for every aquatic insect genome. A lack of aquatic genomes is limiting research progress in the field at both fundamental and applied scales. We argue that the limited availability of aquatic insect genomes is not due to practical limitations—small body sizes or overly complex genomes—but instead reflects a lack of research interest. We call for targeted efforts to expand the availability of aquatic insect genomic resources to empower future research. -

Monte L. Bean Life Science Museum Brigham Young University Provo, Utah 84602 PBRIA a Newsletter for Plecopterologists
No. 10 1990/1991 Monte L. Bean Life Science Museum Brigham Young University Provo, Utah 84602 PBRIA A Newsletter for Plecopterologists EDITORS: Richard W, Baumann Monte L. Bean Life Science Museum Brigham Young University Provo, Utah 84602 Peter Zwick Limnologische Flußstation Max-Planck-Institut für Limnologie, Postfach 260, D-6407, Schlitz, West Germany EDITORIAL ASSISTANT: Bonnie Snow REPORT 3rd N orth A merican Stonefly S ymposium Boris Kondratieff hosted an enthusiastic group of plecopterologists in Fort Collins, Colorado during May 17-19, 1991. More than 30 papers and posters were presented and much fruitful discussion occurred. An enjoyable field trip to the Colorado Rockies took place on Sunday, May 19th, and the weather was excellent. Boris was such a good host that it was difficult to leave, but many participants traveled to Santa Fe, New Mexico to attend the annual meetings of the North American Benthological Society. Bill Stark gave us a way to remember this meeting by producing a T-shirt with a unique “Spirit Fly” design. ANNOUNCEMENT 11th International Stonefly Symposium Stan Szczytko has planned and organized an excellent symposium that will be held at the Tree Haven Biological Station, University of Wisconsin in Tomahawk, Wisconsin, USA. The registration cost of $300 includes lodging, meals, field trip and a T- Shirt. This is a real bargain so hopefully many colleagues and friends will come and participate in the symposium August 17-20, 1992. Stan has promised good weather and good friends even though he will not guarantee that stonefly adults will be collected during the field trip. Printed August 1992 1 OBITUARIES RODNEY L. -

Download .PDF(1340
Stark, Bill P. and Stephen Green. 2011. Eggs of western Nearctic Acroneuriinae (Plecoptera: Perlidae). Illiesia, 7(17):157-166. Available online: http://www2.pms-lj.si/illiesia/Illiesia07-17.pdf EGGS OF WESTERN NEARCTIC ACRONEURIINAE (PLECOPTERA: PERLIDAE) Bill P. Stark1 and Stephen Green2 1,2 Box 4045, Department of Biology, Mississippi College, Clinton, Mississippi, U.S.A. 39058 1 E-mail: [email protected] 2 E-mail: [email protected] ABSTRACT Eggs for western Nearctic acroneuriine species of Calineuria Ricker, Doroneuria Needham & Claassen and Hesperoperla Banks are examined and redescribed based on scanning electron microscopy images taken from specimens collected from a substantial portion of each species range. Within genera, species differences in egg morphology are small and not always useful for species recognition, however eggs from one population of Calineuria are significantly different from those found in other populations and this population is given informal recognition as a possible new species. Keywords: Plecoptera, Calineuria, Doroneuria, Hesperoperla, Egg morphology, Western Nearctic INTRODUCTION occur in the region (Baumann & Olson 1984; Scanning electron microscopy (SEM) is often used Kondratieff & Baumann 2002; Stark 1989; Stark & to elucidate chorionic features for stoneflies (e.g. Gaufin 1976; Stark & Kondratieff 2004; Zuellig et al. Baumann 1973; Grubbs 2005; Isobe 1988; Kondratieff 2006). SEM images for eggs of the primary western 2004; Kondratieff & Kirchner 1996; Nelson 2000; acroneuriine genera, Calineuria Ricker, Doroneuria Sivec & Stark 2002; 2008; Stark & Nelson 1994; Stark Needham & Claassen and Hesperoperla Banks include & Szczytko 1982; 1988; Szczytko & Stewart 1979) and single images for each of these genera in Stark & Nearctic Perlidae were among the earliest stoneflies Gaufin (1976), three images of Hesperoperla hoguei to be studied with this technique (Stark & Gaufin Baumann & Stark (1980) and three images of H. -

Isoperla Bilineata (Group)
Steven R Beaty Biological Assessment Branch North Carolina Division of Water Resources [email protected] 2 DISCLAIMER: This manual is unpublished material. The information contained herein is provisional and is intended only to provide a starting point for the identification of Isoperla within North Carolina. While many of the species treated here can be found in other eastern and southeastern states, caution is advised when attempting to identify Isoperla outside of the study area. Revised and corrected versions are likely to follow. The user assumes all risk and responsibility of taxonomic determinations made in conjunction with this manual. Recommended Citation Beaty, S. R. 2015. A morass of Isoperla nymphs (Plecoptera: Perlodidae) in North Carolina: a photographic guide to their identification. Department of Environment and Natural Resources, Division of Water Resources, Biological Assessment Branch, Raleigh. Nymphs used in this study were reared and associated at the NCDENR Biological Assessment Branch lab (BAB) unless otherwise noted. All photographs in this manual were taken by the Eric Fleek (habitus photos) and Steve Beaty (lacinial photos) unless otherwise noted. They may be used with proper credit. 3 Keys and Literature for eastern Nearctic Isoperla Nymphs Frison, T. H. 1935. The Stoneflies, or Plecoptera, of Illinois. Illinois Natural History Bulletin 20(4): 281-471. • while not containing species of isoperlids that occur in NC, it does contain valuable habitus and mouthpart illustrations of species that are similar to those found in NC (I. bilineata, I richardsoni) Frison, T. H. 1942. Studies of North American Plecoptera with special reference to the fauna of Illinois. Illinois Natural History Bulletin 22(2): 235-355. -

New Records of Stoneflies (Plecoptera) with an Annotated Checklist of the Species for Pennsylvania
The Great Lakes Entomologist Volume 29 Number 3 - Fall 1996 Number 3 - Fall 1996 Article 2 October 1996 New Records of Stoneflies (Plecoptera) With an Annotated Checklist of the Species for Pennsylvania E. C. Masteller Behrend College Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Masteller, E. C. 1996. "New Records of Stoneflies (Plecoptera) With an Annotated Checklist of the Species for Pennsylvania," The Great Lakes Entomologist, vol 29 (3) Available at: https://scholar.valpo.edu/tgle/vol29/iss3/2 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Masteller: New Records of Stoneflies (Plecoptera) With an Annotated Checklis 1996 THE GREAT LAKES ENTOMOlOGIST 107 NEW RECORDS OF STONEFLIES IPLECOPTERA} WITH AN ANNOTATED CHECKLIST OF THE SPECIES FOR PENNSYLVANIA E.C. Masteller1 ABSTRACT Original collections now record 134 species in nine families and 42 gen era. Seventeen new state records include, Allocapnia wrayi, Alloperla cau data, Leuctra maria, Soyedina carolinensis, Tallaperla elisa, Perlesta decipi· ens, P. placida, Neoperla catharae, N. occipitalis, N. stewarti, Cult us decisus decisus, Isoperla francesca, 1. frisoni, 1. lata,1. nana, 1. slossonae, Malirekus hastatus. Five species are removed from the list ofspecies for Pennsylvania. Surdick and Kim (1976) originally recorded 90 species of stoneflies in nine families and 32 genera from Pennsylvania. Since that time, Stark et al. -
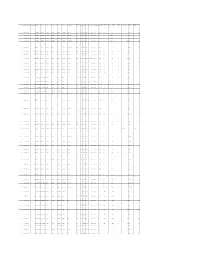
Appendix D: Summary of Accepted ECOTOX Papers
Effect Dur Dur Unit Dur Dur Unit Conc Conc Units Conc Value1 Purity CAS Number Chemical Name Species Number Phylum Class Order Family Genus Species Common Name Group Effect Meas Endpt1 Endpt2 Habitat Plant/Animal Media Orig Orig Preferred Preferred Type Conc Value1 Orig Orig Adjusted 1 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BEH AVO STIM NOAEL terrestrial Animal NONE 30 d 30 d F 2 mg/kg/d 1.96 2 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM LIPD LOAEL aquatic Animal FW 1 h 4.17E-02 d A 4 mg/L 4 3 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM PRCO LOAEL aquatic Animal FW 1 h 4.17E-02 d A 4 mg/L 4 4 298044 Disulfoton 351 Chordata Actinopterygii Perciformes Anabantidae Anabas testudineus Climbing perch BCM BCM LIPD LOAEL aquatic Animal FW 1 h 4.17E-02 d A 10.5 mg/L 10.5 5 298044 Disulfoton 4913 Chordata Mammalia Rodentia Muridae Mus musculus House mouse BCM BCM HXBT LOAEL terrestrial Animal NONE 3 d 3 d F 35.1 uM/kg 35.1 6 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM LOAEL terrestrial Animal NONE 24 h 1 d F 0.26 mg/kg 0.234 7 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM IC50 terrestrial Animal NONE 1 wk 7 d F 0.6 ppm 0.6 8 298044 Disulfoton 4510 Chordata Mammalia Rodentia Muridae Rattus norvegicus Norway rat BCM BCM GBCM NOAEL LOAEL terrestrial Animal NONE -

Ohio EPA Macroinvertebrate Taxonomic Level December 2019 1 Table 1. Current Taxonomic Keys and the Level of Taxonomy Routinely U
Ohio EPA Macroinvertebrate Taxonomic Level December 2019 Table 1. Current taxonomic keys and the level of taxonomy routinely used by the Ohio EPA in streams and rivers for various macroinvertebrate taxonomic classifications. Genera that are reasonably considered to be monotypic in Ohio are also listed. Taxon Subtaxon Taxonomic Level Taxonomic Key(ies) Species Pennak 1989, Thorp & Rogers 2016 Porifera If no gemmules are present identify to family (Spongillidae). Genus Thorp & Rogers 2016 Cnidaria monotypic genera: Cordylophora caspia and Craspedacusta sowerbii Platyhelminthes Class (Turbellaria) Thorp & Rogers 2016 Nemertea Phylum (Nemertea) Thorp & Rogers 2016 Phylum (Nematomorpha) Thorp & Rogers 2016 Nematomorpha Paragordius varius monotypic genus Thorp & Rogers 2016 Genus Thorp & Rogers 2016 Ectoprocta monotypic genera: Cristatella mucedo, Hyalinella punctata, Lophopodella carteri, Paludicella articulata, Pectinatella magnifica, Pottsiella erecta Entoprocta Urnatella gracilis monotypic genus Thorp & Rogers 2016 Polychaeta Class (Polychaeta) Thorp & Rogers 2016 Annelida Oligochaeta Subclass (Oligochaeta) Thorp & Rogers 2016 Hirudinida Species Klemm 1982, Klemm et al. 2015 Anostraca Species Thorp & Rogers 2016 Species (Lynceus Laevicaudata Thorp & Rogers 2016 brachyurus) Spinicaudata Genus Thorp & Rogers 2016 Williams 1972, Thorp & Rogers Isopoda Genus 2016 Holsinger 1972, Thorp & Rogers Amphipoda Genus 2016 Gammaridae: Gammarus Species Holsinger 1972 Crustacea monotypic genera: Apocorophium lacustre, Echinogammarus ischnus, Synurella dentata Species (Taphromysis Mysida Thorp & Rogers 2016 louisianae) Crocker & Barr 1968; Jezerinac 1993, 1995; Jezerinac & Thoma 1984; Taylor 2000; Thoma et al. Cambaridae Species 2005; Thoma & Stocker 2009; Crandall & De Grave 2017; Glon et al. 2018 Species (Palaemon Pennak 1989, Palaemonidae kadiakensis) Thorp & Rogers 2016 1 Ohio EPA Macroinvertebrate Taxonomic Level December 2019 Taxon Subtaxon Taxonomic Level Taxonomic Key(ies) Informal grouping of the Arachnida Hydrachnidia Smith 2001 water mites Genus Morse et al. -

Life History and Production of Mayflies, Stoneflies, and Caddisflies (Ephemeroptera, Plecoptera, and Trichoptera) in a Spring-Fe
Color profile: Generic CMYK printer profile Composite Default screen 1083 Life history and production of mayflies, stoneflies, and caddisflies (Ephemeroptera, Plecoptera, and Trichoptera) in a spring-fed stream in Prince Edward Island, Canada: evidence for population asynchrony in spring habitats? Michelle Dobrin and Donna J. Giberson Abstract: We examined the life history and production of the Ephemeroptera, Plecoptera, and Trichoptera (EPT) commu- nity along a 500-m stretch of a hydrologically stable cold springbrook in Prince Edward Island during 1997 and 1998. Six mayfly species (Ephemeroptera), 6 stonefly species (Plecoptera), and 11 caddisfly species (Trichoptera) were collected from benthic and emergence samples from five sites in Balsam Hollow Brook. Eleven species were abundant enough for life-history and production analysis: Baetis tricaudatus, Cinygmula subaequalis, Epeorus (Iron) fragilis,andEpeorus (Iron) pleuralis (Ephemeroptera), Paracapnia angulata, Sweltsa naica, Leuctra ferruginea, Amphinemura nigritta,and Nemoura trispinosa (Plecoptera), and Parapsyche apicalis and Rhyacophila brunnea (Trichoptera). Life-cycle timing of EPT taxa in Balsam Hollow Brook was generally similar to other literature reports, but several species showed extended emergence periods when compared with other studies, suggesting a reduction in synchronization of life-cycle timing, pos- sibly as a result of the thermal patterns in the stream. Total EPT secondary production (June 1997 to May 1998) was 2.74–2.80 g·m–2·year–1 dry mass (size-frequency method). Mayflies were dominant, with a production rate of 2.2 g·m–2·year–1 dry mass, followed by caddisflies at 0.41 g·m–2·year–1 dry mass, and stoneflies at 0.19 g·m–2·year–1 dry mass. -

About the Book the Format Acknowledgments
About the Book For more than ten years I have been working on a book on bryophyte ecology and was joined by Heinjo During, who has been very helpful in critiquing multiple versions of the chapters. But as the book progressed, the field of bryophyte ecology progressed faster. No chapter ever seemed to stay finished, hence the decision to publish online. Furthermore, rather than being a textbook, it is evolving into an encyclopedia that would be at least three volumes. Having reached the age when I could retire whenever I wanted to, I no longer needed be so concerned with the publish or perish paradigm. In keeping with the sharing nature of bryologists, and the need to educate the non-bryologists about the nature and role of bryophytes in the ecosystem, it seemed my personal goals could best be accomplished by publishing online. This has several advantages for me. I can choose the format I want, I can include lots of color images, and I can post chapters or parts of chapters as I complete them and update later if I find it important. Throughout the book I have posed questions. I have even attempt to offer hypotheses for many of these. It is my hope that these questions and hypotheses will inspire students of all ages to attempt to answer these. Some are simple and could even be done by elementary school children. Others are suitable for undergraduate projects. And some will take lifelong work or a large team of researchers around the world. Have fun with them! The Format The decision to publish Bryophyte Ecology as an ebook occurred after I had a publisher, and I am sure I have not thought of all the complexities of publishing as I complete things, rather than in the order of the planned organization. -

A Review of the Stoneflies of the Rock River, Illinois
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Illinois Digital Environment for Access to Learning and Scholarship Repository ILLINOI S UNIVERSITY OF ILLINOIS AT URBANA-CHAMPAIGN PRODUCTION NOTE University of Illinois at Urbana-Champaign Library Large-scale Digitization Project, 2007. A REVIEW OF THE STONEFLIES OF THE ROCK RIVER, ILLINOIS Dr. Donald W. Webb Center For Biodiversity Illinois Natural History Survey 607 East Peabody Drive Champaign, Illinois 61820 TECHNICAL REPORT 2002 (11) ILLINOIS NATURAL HISTORY SURVEY CENTER FOR BIODIVERSITY PREPARED FOR Division of Natural Heritage Office of Resource Conservation Illinois Department of Natural Resources One Natural Resources Way Springfield, IL 62702 Abstract During the 1990's, collecting was done along the Rock River in a effort to collect winter stoneflies (those species emerging from December through March). In 1997, collecting was done in and around Rock Island in an effort to collect Alloperla roberti. During April, May, and June of 2002, collecting for spring emerging stoneflies was conducted at nine sites along the Rock River from Rock Island to Rockton. Historically, 25 species of stoneflies (Insecta: Plecoptera) have been reported from the Rock River. Based on collecting from 1990-2002 eleven species (Acroneuria abnormis, Allocapnia granulata,Allocapnia vivipara Isoperla bilineata, Isoperla richardsoni,Perlesta golconda, Perlesta decipiens, Perlinella ephyre, Pteronarcys pictetii, Taeniopteryx burksi, Taeniopteryx nivalis) remain established within the Rock River. Acroneuria abnormis was previously very abundant along the length of the Rock River, but now is considered very rare. Allocapnia vivipara, the most common species of stonefly in Illinois and primarily a small stream species, appears to have been replaced by Allocapnia granulata in the Rock River. -

Plecoptera: Perlidae), with an Annotated Checklist of the Subfamily in the Realm
Opusc. Zool. Budapest, 2016, 47(2): 173–196 On the identity of some Oriental Acroneuriinae taxa (Plecoptera: Perlidae), with an annotated checklist of the subfamily in the realm D. MURÁNYI1 & W.H. LI2 1Dávid Murányi, Department of Civil and Environmental Engineering, Ehime University, Bunkyo-cho 3, Matsuyama, 790-8577 Japan, and Department of Zoology, Hungarian Natural History Museum, H-1088 Budapest, Baross u. 13, Hungary. E-mail: [email protected], [email protected] 2Weihai Li, Department of Plant Protection, Henan Institute of Science and Technology, Xinxiang, Henan, 453003 China. E-mail: [email protected] Abstract. The monotypic Taiwanese genus Mesoperla Klapálek, 1913 is redescribed on the basis of a male syntype specimen, and its affinities are re-evaluated. The single female type specimen of further two Oriental monotypic genera, Kalidasia Klapálek, 1914 and Nirvania Klapálek, 1914, are confirmed to be lost or destroyed respectively; both genera are considered as nomina dubia. The Sichuan endemic Acroneuria grahami Wu & Claassen, 1934 is redescribed on the basis of male holotype. Distinctive characters of the genus Brahmana Klapálek, 1914 consisting of five, inadequately known Oriental species are discussed. Flavoperla needhami (Klapálek, 1916) and Sinacroneuria sinica (Yang & Yang, 1998) comb. novae are suggested for an Indian species originally described in Gibosia Okamoto, 1912 and a Chinese species originally described in Acroneuria Pictet, 1841. At present, 62 species of Acroneuriinae, classified in 10 valid genera are reported from the Oriental Realm but 29 species are inadequately known. A key is presented to distinguish males of the Asian Acroneuriinae genera. Asian distribution of each genera are detailed and depicted on a map. -

Nymphs of North American Perlodinae Genera (Plecoptera: Perlodidae)
Great Basin Naturalist Volume 44 Number 3 Article 1 7-31-1984 Nymphs of North American Perlodinae genera (Plecoptera: Perlodidae) Kenneth W. Stewart North Texas State University, Denton, Texas Bill P. Stark Mississippi College, Clinton, Mississippi Follow this and additional works at: https://scholarsarchive.byu.edu/gbn Recommended Citation Stewart, Kenneth W. and Stark, Bill P. (1984) "Nymphs of North American Perlodinae genera (Plecoptera: Perlodidae)," Great Basin Naturalist: Vol. 44 : No. 3 , Article 1. Available at: https://scholarsarchive.byu.edu/gbn/vol44/iss3/1 This Article is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Great Basin Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. The Great Basin Naturalist Published at Provo, Utah, by Brigham Young University ISSN 0017-3614 Volume 44 July 31, 1984 No. 3 NYMPHS OF NORTH AMERICAN PERLODINAE GENERA (PLECOPTERA: PERLODIDAE)' Kenneth VV. Stewart- and Bill P. Stark' Abstract.— Nymphs of the type or other representative species of the 22 North American Perlodinae genera are comparatively described and illustrated for the first time. The first complete generic key for the subfamily incorporates recent nymph discoveries and revisions in classification. References to all previous nymph descriptions and illustrations and major life cycle and food habits studies are given for the 53 North American species in the subfamilv, and a listing of species and their current distributions by states and provinces is provided for each genus. The previously unknown nymph of Chcrnokrihts misnomus is described and illustrated.