Journal of Hazardous Materials 297 (2015) 17–24
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Teratogenicity and Embryotoxicity of Organophosphorus Compounds in Animal Models - a Short Review
Mil. Med. Sci. Lett. (Voj. Zdrav. Listy) 2012, vol. 81(1), p. 16-26 ISSN 0372-7025 DOI: 10.31482/mmsl.2012.003 REVIEW ARTICLE TERATOGENICITY AND EMBRYOTOXICITY OF ORGANOPHOSPHORUS COMPOUNDS IN ANIMAL MODELS - A SHORT REVIEW Syed M Nurulain and M Shafiullah Department of Pharmacology and Therapeutic, Faculty of Medicine and Health Sciences, UAE University, AlAin, UAE. P.O.Box 17666, Alain, UAE Received 9 th December 2011. Revised 12 th February 2012. Published 9 th March 2012. Summary Organophosphorus compounds (OPCs) are a wide group of compounds both structurally and functionally. Each OPC has a unique toxicological profile. The exposure to this type of poison is not limited only to certain occupationally exposed people but also to children, women, pregnant women; all have chances to be exposed to this poison. During the recent past years it has been reported in many poison epidemiological studies and case reports that exposure of OPCs during pregnancy caused malformed fetuses, neural tube defect (NTD) and shortening of pregnancy. The literature for animal models reveals inconclusive evidence. The generalized view is that they are neither teratogenic nor embryotoxic. But it is not true. There is a lack of systematic study and scarcity of reports on the topic. The present study was undertaken to investigate the teratogenicity induced by organophosphorus compounds in different animal models by literature review. Literature was searched by Toxicology Data NetWork (TOXNET), Developmental and Reproductive Toxicology Database (DART), Toxicology Literature Online (TOXLINE), Hazardous Substances Data Bank (HSDB), Pubmed Central, Entrez-Pubmed, Science Direct, Directory Of Open Access Journal (DOAJ), Google Scholar and International Program on Chemical Safety (IPCS-INCHEM), Embase. -

Determination of Age-Related Differences in Activation and Detoxication of Organophosphates in Rat and Human Tissues
Mississippi State University Scholars Junction Theses and Dissertations Theses and Dissertations 8-10-2018 Determination of Age-Related Differences in Activation and Detoxication of Organophosphates in Rat and Human Tissues Edward Caldwell Meek Follow this and additional works at: https://scholarsjunction.msstate.edu/td Recommended Citation Meek, Edward Caldwell, "Determination of Age-Related Differences in Activation and Detoxication of Organophosphates in Rat and Human Tissues" (2018). Theses and Dissertations. 1339. https://scholarsjunction.msstate.edu/td/1339 This Dissertation - Open Access is brought to you for free and open access by the Theses and Dissertations at Scholars Junction. It has been accepted for inclusion in Theses and Dissertations by an authorized administrator of Scholars Junction. For more information, please contact [email protected]. Template A v3.0 (beta): Created by J. Nail 06/2015 Determination of age-related differences in activation and detoxication of organophosphates in rat and human tissues By TITLE PAGE Edward Caldwell Meek A Dissertation Submitted to the Faculty of Mississippi State University in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy in Environmental Toxicology in the College of Veterinary Medicine Mississippi State, Mississippi August 2018 Copyright by COPYRIGHT PAGE Edward Caldwell Meek 2018 Determination of age-related differences in activation and detoxication of organophosphates in rat and human tissues By APPROVAL PAGE Edward Caldwell Meek -

20110628140658587.Pdf
1210 YUE ET AL.:JOURNAL OF AOAC INTERNATIONAL VOL. 91, NO. 5, 2008 SPECIAL GUEST EDITOR SECTION High-Performance Thin-Layer Chromatographic Analysis of Selected Organophosphorous Pesticide Residues in Tea YONGDE YUE International Center for Bamboo and Rattan, 100102, Beijing, People’s Republic of China RONG ZHANG and WEI FAN Key Laboratory for Agri-Food Safety of Anhui Province, School of Resources and Environment, Anhui Agricultural University, Hefei, Anhui, 230036, People’s Republic of China FENG TANG International Center for Bamboo and Rattan, 100102, Beijing, People’s Republic of China The separation of 9 organophosphates determination procedure. In many cases, searching for a fast (monocrotophos, quinalphos, triazophos, and efficient method for enrichment and determination of parathion-methyl, isofenphos-methyl, temephos, organophosphorous residues from complex matrixes is still a parathion, phoxim-ethyl, and chlorpyrifos) by challenge for many researchers. high-performance thin-layer chromatography In many analytical situations, modern quantitative (HPTLC) with automated multiple development was high-performance thin-layer chromatography (HPTLC), studied. The HPTLC method was developed and when properly performed by well-trained analysts, has many validated for analysis of residues of phoxim-ethyl advantages over HPLC or GC. These advantages include and chlorpyrifos in tea. The sample was extracted simplicity of operation, the availability of many sensitive and with acetonitrile and cleaned up by ENVI-CARB selective reagents for detection and confirmation without solid-phase extraction. The extract was directly interference from the mobile phase, the ability to repeat applied as bands to glass-backed silica gel 60F254 detection and quantification at any time with changed HPTLC plates. -

Triazofos 60% LC
Excel Crop Care Limited, India Material Safety Data Sheet Triazofos 60% LC 1. Chemical Product and Company Identification Product Name : Triazofos 60% LC Formula : C12 H16N3O3PS Molecuar Weight : 313.3 : O,O-diethyl O-(1-phenyl-1H-1,2,4-triazol-3-yl) phosphorothioate. Chemical Name Manufacturer : Excel Crop Care Limited 184 / 87, S. V. Road, Jogeshwari (West), Mumbai, (India). Pin code: 400 102 Contact no: +912266464200 Emergency Contact Details : +91 22 42522201 : +91 22 42522202 : +91 278 2212320 2. Composition/Information on Ingredients Component CAS No. % W/W content Triazofos 24017-47-8 60 minimum Xylene 1330-20-7 Q. S. 3. Hazard Identification Emergency Overview Appearance and odour : Pale yellow to dark brown clear liquid, with typical phosphate ester odour Warning Statements : Keep out of reach of children Potential Health Effects Likely Routs of Exposure : Skin contact, Inhalation and Ingestion Eye Contact :Unlikely to cause irritation to eye Skin contact : Not irritating to skin Ingestion :If swallowed, this substance is considered to be toxic and likely to Cause damages to internal organ Version: ECCL/MSDS/Triazo60LC/1 © Excel Crop Care Ltd Page 1 of 7 DoC: Sept 14, 2014 Private Document DoU: Sept 25, 2014 Issue Date: Oct 1, 2014 Excel Crop Care Limited, India Material Safety Data Sheet Triazofos 60% LC Inhalation : Repeated exposure may cause cholinesterase inhibition. Environmental Hazards : The product is an organophosphate insecticide 4. First aid measures and Antidote Emergency and First Aid Procedure – If in Eyes : Immediately flush with plenty of clean water for 15-20 minutes. Remove contact lenses if present after 5 minutes of washing. -

Sister Chromatid Exchanges Induced by the Organophosphorus Insecticides Methyl Parathion, Dimethoate, Phoxim and Methyl Azinfos in Cultured Human Lymphocytes
Contam. Arnbient. 3. 63-70. 1981 SISTER CHROMATID EXCHANGES INDUCED BY THE ORGANOPHOSPHORUS INSECTICIDES METHYL PARATHION, DIMETHOATE, PHOXIM AND METHYL AZINFOS IN CULTURED HUMAN LYMPHOCYTES SANDRAGOMEZ-ARROYO, NORMANORIEGA-ALDANA, DOLORESJUAREZ-RODRI- GUEZ and RAFAELVILLALOBOS-PIETRINI Laboratorio de Citogenética y Mutagénesis Ambientales, Centro de Ciencias de la Atmós- fera, Universidad Nacional Autánoma de México, Coyoacán 04510, México, D.F. México (Recibido septiembre 1987, Aceptado diciembre 1987) ABSTRACT The cytogenetic effect of the organophosphorus insecticides methyl parathion or folidol. dimethoate or rogor. phoxirn or bay 77488 and methyl azinfos or gusathion was evahiated by means of the productions of sister chromatid exchanges (SCE) in cultured human lymphocytes. Positive results were obtained with methyl parathion, dimethoate and phoxim, while a negative response was achieved with methyl azinfos. The highest con- centrations of all substances decreased the SCE. possibly due to their toxicity, produ- cing the death of the damaged cells and therefore eiiminating the SCE expression. RESUMEN Se evaluó el afecto citogenbtico de los insecticidas organofosforados metil paration o foiidol, dimetoato o rogor. foxim o bay 77488 y metil azinfos o gusation mediante el anáiisis de intercambio de cromátidas hermanas (ICH) en cultivo de linfocitos humanos. Se obtuvieron resultados positivos con metil paratión, dimetoato y foxim, en tanto que con metil azinfos la respuesta fue negativa. A las concentraciones más altas de todas las sustancias empleadas disminuyeron los ICH, debido posiblemente a su elevada to- xicidad, provocando la muerte de las células más dañadas e impidiendo de esta manera la expresión del ICH. INTRODUCTION The organophosphorus insecticides appear for the first time in 1945 in Gerrnany. -
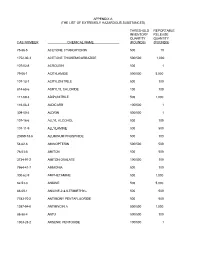
The List of Extremely Hazardous Substances)
APPENDIX A (THE LIST OF EXTREMELY HAZARDOUS SUBSTANCES) THRESHOLD REPORTABLE INVENTORY RELEASE QUANTITY QUANTITY CAS NUMBER CHEMICAL NAME (POUNDS) (POUNDS) 75-86-5 ACETONE CYANOHYDRIN 500 10 1752-30-3 ACETONE THIOSEMICARBAZIDE 500/500 1,000 107-02-8 ACROLEIN 500 1 79-06-1 ACRYLAMIDE 500/500 5,000 107-13-1 ACRYLONITRILE 500 100 814-68-6 ACRYLYL CHLORIDE 100 100 111-69-3 ADIPONITRILE 500 1,000 116-06-3 ALDICARB 100/500 1 309-00-2 ALDRIN 500/500 1 107-18-6 ALLYL ALCOHOL 500 100 107-11-9 ALLYLAMINE 500 500 20859-73-8 ALUMINUM PHOSPHIDE 500 100 54-62-6 AMINOPTERIN 500/500 500 78-53-5 AMITON 500 500 3734-97-2 AMITON OXALATE 100/500 100 7664-41-7 AMMONIA 500 100 300-62-9 AMPHETAMINE 500 1,000 62-53-3 ANILINE 500 5,000 88-05-1 ANILINE,2,4,6-TRIMETHYL- 500 500 7783-70-2 ANTIMONY PENTAFLUORIDE 500 500 1397-94-0 ANTIMYCIN A 500/500 1,000 86-88-4 ANTU 500/500 100 1303-28-2 ARSENIC PENTOXIDE 100/500 1 THRESHOLD REPORTABLE INVENTORY RELEASE QUANTITY QUANTITY CAS NUMBER CHEMICAL NAME (POUNDS) (POUNDS) 1327-53-3 ARSENOUS OXIDE 100/500 1 7784-34-1 ARSENOUS TRICHLORIDE 500 1 7784-42-1 ARSINE 100 100 2642-71-9 AZINPHOS-ETHYL 100/500 100 86-50-0 AZINPHOS-METHYL 10/500 1 98-87-3 BENZAL CHLORIDE 500 5,000 98-16-8 BENZENAMINE, 3-(TRIFLUOROMETHYL)- 500 500 100-14-1 BENZENE, 1-(CHLOROMETHYL)-4-NITRO- 500/500 500 98-05-5 BENZENEARSONIC ACID 10/500 10 3615-21-2 BENZIMIDAZOLE, 4,5-DICHLORO-2-(TRI- 500/500 500 FLUOROMETHYL)- 98-07-7 BENZOTRICHLORIDE 100 10 100-44-7 BENZYL CHLORIDE 500 100 140-29-4 BENZYL CYANIDE 500 500 15271-41-7 BICYCLO[2.2.1]HEPTANE-2-CARBONITRILE,5- -

Application of Dried Blood Spots for the Determination of Organophosphorus Pesticides by GC-MS/MS
UNIVERSIDADE DA BEIRA INTERIOR Ciências Application of dried blood spots for the determination of organophosphorus pesticides by GC-MS/MS Sofia Pires Seixo Soares Dissertação para obtenção do Grau de Mestre em Bioquímica (2º ciclo de estudos) Orientador: Doutor Mário Jorge Dinis Barroso Co-orientador: Professora Doutora Maria Eugenia Gallardo Alba Covilhã, junho de 2018 Agradecimentos A realização de uma dissertação de mestrado é o culminar de muitas horas de estudo, reflexão e trabalho pessoal, mas representa também uma etapa que não seria possível sem o contributo fundamental de várias pessoas. À Doutora Eugénia Gallardo e ao Doutor Mário Barroso, como excelentes mentores que são, quero agradecer toda a orientação, motivação, incentivo e confiança. Toda a ajuda e partilha de experiência e conhecimento durante todo este percurso. Ao Tiago Rosado, pela imprescindível ajuda e disponibilidade constante, bem como pela amizade. À Joana Gonçalves e ao Ângelo Luís um agradecimento especial pela amizade e carinho demonstrado ao longo deste ano. A todos, quero agradecer pelo bom ambiente proporcionado. Aos meus pais Ana Soares e José Soares, a quem dedico este feito, por me darem a possibilidade de concretizar conquistas como esta, pelo apoio incondicional, por sempre terem acreditado em mim e por fazerem parte deste momento. Aos meus avós e restante família, por todo o apoio e preocupação. Um agradecimento particular à Inês Ramos, à Rita Marques, à Maria Cunha e ao Pedro Batista pelo companheirismo, amizade e apoio de há tantos anos. E por último, mas nunca menos importante, um enorme obrigada ao meu namorado António Fernandes por toda a paciência, pelo incansável apoio, carinho e ajuda. -

Redalyc.Luminescence Based Detection of Trinitrophenol And
Journal of the Mexican Chemical Society ISSN: 1870-249X [email protected] Sociedad Química de México México Kaur, Rupinder; Chhibber, Manmohan; Mahata, Partha; Mittal, Susheel K. Luminescence Based Detection of Trinitrophenol and Aromatic Organophosphorous Pesticides Using a Coordination Polymer Journal of the Mexican Chemical Society, vol. 61, núm. 4, 2017, pp. 336-341 Sociedad Química de México Distrito Federal, México Available in: http://www.redalyc.org/articulo.oa?id=47554335008 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative J. Mex. Chem. Soc. 2017, 61(4),336-341 Article © 2017, Sociedad Química de México ISSN 1870-249X Luminescence Based Detection of Trinitrophenol and Aromatic Organophosphorous Pesticides Using a Coordination Polymer Rupinder Kaur,1 Manmohan Chhibber,1 Partha Mahata2 and Susheel K. Mittal*1 1 School of Chemistry & Biochemistry, Thapar University, Patiala-147004, India. Email: [email protected] 2 Department of Chemistry, Jadavpur University, Kolkata-700 032, India Email: [email protected] Received February 3rd, 2017; Accepted September 18th, 2017 Abstract: The fluorescent properties of a coordination polymer Resumen: Las propiedades fluorescentes del polímero de coordinación (CP), 1, were used as turn-on and turn-off detector for nitroaromatics (CP), 1, fueron utilizadas -

Investigation on the Behavior of Pesticides in Atmosphere
Aerosol and Air Quality Research, 11: 783–790, 2011 Copyright © Taiwan Association for Aerosol Research ISSN: 1680-8584 print / 2071-1409 online doi: 10.4209/aaqr.2010.10.0085 Investigation on the Behavior of Pesticides in Atmosphere Pasquale Avino1, Giuseppe Cinelli2, Ivan Notardonato2, Mario V. Russo2* 1 DIPIA, INAIL (ex-ISPESL), via Urbana 167, 00184 Rome, Italy 2 Faculty of Agriculture, University of Molise, via De Sanctis, Campobasso, Italy ABSTRACT Although pesticides are widely used in agriculture, they and in particular the relative residues in foodstuffs, water and atmosphere, may cause remarkable sanitary problems due to the harmful effects (carcinogenic and mutagenic effects) on the human health. In fact, their spread in waters and atmosphere can produce undesired effects on various organisms and/or water contamination. This paper shows an analytical approach based on XAD-2 adsorbent and GC analysis for evaluating the pesticide trend in atmosphere: in particular, the pesticides investigated are omethoate, dicrotofos, disulfoton, dimethoate, parathion methyl, formothion, paraoxon ethyl, malaoxon, parathion ethyl, iodofenfos and triazofos. For the analytical methodology a linearity response was obtained (r2 = 0.9988) in GC-NPD whereas the limits of detection range between 2 and 5 pg/μL in GC-NPD with a Relative Standard Deviation below 9.5. Finally, this approach has been successfully applied to real samples: the results show that dimethoate concentration decreases with increasing distance from the sampling site but it is still persistent in atmosphere after few days from the pesticide spraying. Keywords: Pesticides; XAD-2 adsorbent; GC analysis; Atmosphere; Air quality. INTRODUCTION 2009). One of the most important OP reactions is water hydrolysis. -

Environmental Health Criteria 63 ORGANOPHOSPHORUS
Environmental Health Criteria 63 ORGANOPHOSPHORUS INSECTICIDES: A GENERAL INTRODUCTION Please note that the layout and pagination of this web version are not identical with the printed version. Organophophorus insecticides: a general introduction (EHC 63, 1986) INTERNATIONAL PROGRAMME ON CHEMICAL SAFETY ENVIRONMENTAL HEALTH CRITERIA 63 ORGANOPHOSPHORUS INSECTICIDES: A GENERAL INTRODUCTION This report contains the collective views of an international group of experts and does not necessarily represent the decisions or the stated policy of the United Nations Environment Programme, the International Labour Organisation, or the World Health Organization. Published under the joint sponsorship of the United Nations Environment Programme, the International Labour Organisation, and the World Health Organization World Health Orgnization Geneva, 1986 The International Programme on Chemical Safety (IPCS) is a joint venture of the United Nations Environment Programme, the International Labour Organisation, and the World Health Organization. The main objective of the IPCS is to carry out and disseminate evaluations of the effects of chemicals on human health and the quality of the environment. Supporting activities include the development of epidemiological, experimental laboratory, and risk-assessment methods that could produce internationally comparable results, and the development of manpower in the field of toxicology. Other activities carried out by the IPCS include the development of know-how for coping with chemical accidents, coordination -
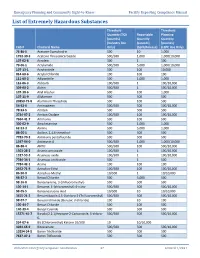
List of Extremely Hazardous Substances
Emergency Planning and Community Right-to-Know Facility Reporting Compliance Manual List of Extremely Hazardous Substances Threshold Threshold Quantity (TQ) Reportable Planning (pounds) Quantity Quantity (Industry Use (pounds) (pounds) CAS # Chemical Name Only) (Spill/Release) (LEPC Use Only) 75-86-5 Acetone Cyanohydrin 500 10 1,000 1752-30-3 Acetone Thiosemicarbazide 500/500 1,000 1,000/10,000 107-02-8 Acrolein 500 1 500 79-06-1 Acrylamide 500/500 5,000 1,000/10,000 107-13-1 Acrylonitrile 500 100 10,000 814-68-6 Acrylyl Chloride 100 100 100 111-69-3 Adiponitrile 500 1,000 1,000 116-06-3 Aldicarb 100/500 1 100/10,000 309-00-2 Aldrin 500/500 1 500/10,000 107-18-6 Allyl Alcohol 500 100 1,000 107-11-9 Allylamine 500 500 500 20859-73-8 Aluminum Phosphide 500 100 500 54-62-6 Aminopterin 500/500 500 500/10,000 78-53-5 Amiton 500 500 500 3734-97-2 Amiton Oxalate 100/500 100 100/10,000 7664-41-7 Ammonia 500 100 500 300-62-9 Amphetamine 500 1,000 1,000 62-53-3 Aniline 500 5,000 1,000 88-05-1 Aniline, 2,4,6-trimethyl- 500 500 500 7783-70-2 Antimony pentafluoride 500 500 500 1397-94-0 Antimycin A 500/500 1,000 1,000/10,000 86-88-4 ANTU 500/500 100 500/10,000 1303-28-2 Arsenic pentoxide 100/500 1 100/10,000 1327-53-3 Arsenous oxide 100/500 1 100/10,000 7784-34-1 Arsenous trichloride 500 1 500 7784-42-1 Arsine 100 100 100 2642-71-9 Azinphos-Ethyl 100/500 100 100/10,000 86-50-0 Azinphos-Methyl 10/500 1 10/10,000 98-87-3 Benzal Chloride 500 5,000 500 98-16-8 Benzenamine, 3-(trifluoromethyl)- 500 500 500 100-14-1 Benzene, 1-(chloromethyl)-4-nitro- 500/500 -

Recommended Classification of Pesticides by Hazard and Guidelines to Classification 2019 Theinternational Programme on Chemical Safety (IPCS) Was Established in 1980
The WHO Recommended Classi cation of Pesticides by Hazard and Guidelines to Classi cation 2019 cation Hazard of Pesticides by and Guidelines to Classi The WHO Recommended Classi The WHO Recommended Classi cation of Pesticides by Hazard and Guidelines to Classi cation 2019 The WHO Recommended Classification of Pesticides by Hazard and Guidelines to Classification 2019 TheInternational Programme on Chemical Safety (IPCS) was established in 1980. The overall objectives of the IPCS are to establish the scientific basis for assessment of the risk to human health and the environment from exposure to chemicals, through international peer review processes, as a prerequisite for the promotion of chemical safety, and to provide technical assistance in strengthening national capacities for the sound management of chemicals. This publication was developed in the IOMC context. The contents do not necessarily reflect the views or stated policies of individual IOMC Participating Organizations. The Inter-Organization Programme for the Sound Management of Chemicals (IOMC) was established in 1995 following recommendations made by the 1992 UN Conference on Environment and Development to strengthen cooperation and increase international coordination in the field of chemical safety. The Participating Organizations are: FAO, ILO, UNDP, UNEP, UNIDO, UNITAR, WHO, World Bank and OECD. The purpose of the IOMC is to promote coordination of the policies and activities pursued by the Participating Organizations, jointly or separately, to achieve the sound management of chemicals in relation to human health and the environment. WHO recommended classification of pesticides by hazard and guidelines to classification, 2019 edition ISBN 978-92-4-000566-2 (electronic version) ISBN 978-92-4-000567-9 (print version) ISSN 1684-1042 © World Health Organization 2020 Some rights reserved.