Scientific Information on a Compilation of Nabugabo Ramsar Site, Uganda
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Impact of Climate Change at Lake Victoria
East Africa Living Lakes Network, C/O OSIENALA (Friends of Lake Victoria). Dunga Beach Kisumu Ann Nabangala Obae. Coordinator. Phone: +254-20-3588681. Email: [email protected] Background It is the Second largest fresh water lake in the World and it is surrounded by three East African States: With 6% in Kenya; Tanzania 52% and Uganda 42%. It is Located at 0:21 0 North and 3:00 0 South of the Equator Lake Victoria has a total length of 3,440 kms and 240 kms wide from East to West and is 1,134 meters above sea level with maximum depth of 82m.Its surface area is 68,870 km 2, catchment area of 180,950 km 2 Generally shallow with maximum depth of 84 meters and mean depth of 40 meters Average inflows and out flows of Lake Victoria Type of flow Flow (m3/s) Percentage (%) Inflows Rain over Lake 3,631 82 Basin Discharge 778 18 Type of flow Flow ( m3/s) Percentage (%) Out flows Evaporation from lake -3,300 76 Nile River - 1,046 24 Balance +33 Sources of Lake Victoria from Kenyan water towers Sondu Miriu river Yala river Nzoia river Mara River Kuja river Impacts and effects Changes in water budget are respectively accompanied by water level fluctuation and promote thermal structures which result in nutrient and food web dynamics. Studies have proved that there is a positive correlation between water level and fish landings (Williams, l972) The abundant fish catches are highly correlated with rainfall and lake levels. The records show that the catches reduced to between 60% and 70% during the current reduction of water level in Nyanza Gulf. -

Lake Opeta Wetland System Ramsar Information Sheet
Information Sheet on Ramsar Wetlands (RIS) Categories approved by Recommendation 4.7, as amended by Resolution VIII.13 of the Conference of the Contracting Parties. Note for compilers: 1. The RIS should be completed in accordance with the attached Explanatory Notes and Guidelines for completing the Information Sheet on Ramsar Wetlands. Compilers are strongly advised to read this guidance before filling in the RIS. 2. Once completed, the RIS (and accompanying map(s)) should be submitted to the Ramsar Bureau. Compilers are strongly urged to provide an electronic (MS Word) copy of the RIS and, where possible, digital copies of maps. LAKE OPETA WETLAND SYSTEM RAMSAR INFORMATION SHEET 1. Name and address of the RIS Compiler: Achilles Byaruhanga and Stephen Kigoolo NatureUganda Plot 83, Tufnel Drive, Kamwokya P. O Box 27034, Kampala - Uganda. Tel: 256 41 540719 Fax no: 256 41 533 528 E-mail: [email protected] 2. Date: 17 September 2005. 3. Country: The Republic of Uganda 4. Name of the Ramsar site: Lake Opeta Wetland System 5. Map of the Ramsar site: Hard copy: attached Digital (electronic) format: yes 6. Geographical coordinates: 34004" - 34024" E and 01033" - 01051" N,. 7. General Location: Lake Opeta and its surrounding swamps is located in north eastern Uganda, in four districts of Nakapiripirit (Namalu sub-county and Lorachat sub-county), Sironko (Bunambutye sub- county), Katakwi (Usuku county, Magoro sub-county) and Kumi (Kolir sub-county, Malera sub-county and Ongino sub-county). It is located 25 kms from Kumi town and the northern part borders a Wildlife Reserve, Pian-Upe Wildlife Reserve and Teso Community Reserve. -

"A Revision of the Freshwater Crabs of Lake Kivu, East Africa."
Northern Michigan University NMU Commons Journal Articles FacWorks 2011 "A revision of the freshwater crabs of Lake Kivu, East Africa." Neil Cumberlidge Northern Michigan University Kirstin S. Meyer Follow this and additional works at: https://commons.nmu.edu/facwork_journalarticles Part of the Biology Commons Recommended Citation Cumberlidge, Neil and Meyer, Kirstin S., " "A revision of the freshwater crabs of Lake Kivu, East Africa." " (2011). Journal Articles. 30. https://commons.nmu.edu/facwork_journalarticles/30 This Journal Article is brought to you for free and open access by the FacWorks at NMU Commons. It has been accepted for inclusion in Journal Articles by an authorized administrator of NMU Commons. For more information, please contact [email protected],[email protected]. This article was downloaded by: [Cumberlidge, Neil] On: 16 June 2011 Access details: Access Details: [subscription number 938476138] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Journal of Natural History Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713192031 The freshwater crabs of Lake Kivu (Crustacea: Decapoda: Brachyura: Potamonautidae) Neil Cumberlidgea; Kirstin S. Meyera a Department of Biology, Northern Michigan University, Marquette, Michigan, USA Online publication date: 08 June 2011 To cite this Article Cumberlidge, Neil and Meyer, Kirstin S.(2011) 'The freshwater crabs of Lake Kivu (Crustacea: Decapoda: Brachyura: Potamonautidae)', Journal of Natural History, 45: 29, 1835 — 1857 To link to this Article: DOI: 10.1080/00222933.2011.562618 URL: http://dx.doi.org/10.1080/00222933.2011.562618 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. -

Water Resources of Uganda: an Assessment and Review
Journal of Water Resource and Protection, 2014, 6, 1297-1315 Published Online October 2014 in SciRes. http://www.scirp.org/journal/jwarp http://dx.doi.org/10.4236/jwarp.2014.614120 Water Resources of Uganda: An Assessment and Review Francis N. W. Nsubuga1,2*, Edith N. Namutebi3, Masoud Nsubuga-Ssenfuma2 1Department of Geography, Geoinformatics and Meteorology, University of Pretoria, Pretoria, South Africa 2National Environmental Consult Ltd., Kampala, Uganda 3Ministry of Foreign Affairs, Kampala, Uganda Email: *[email protected] Received 1 August 2014; revised 26 August 2014; accepted 18 September 2014 Copyright © 2014 by authors and Scientific Research Publishing Inc. This work is licensed under the Creative Commons Attribution International License (CC BY). http://creativecommons.org/licenses/by/4.0/ Abstract Water resources of a country constitute one of its vital assets that significantly contribute to the socio-economic development and poverty eradication. However, this resource is unevenly distri- buted in both time and space. The major source of water for these resources is direct rainfall, which is recently experiencing variability that threatens the distribution of resources and water availability in Uganda. The annual rainfall received in Uganda varies from 500 mm to 2800 mm, with an average of 1180 mm received in two main seasons. The spatial distribution of rainfall has resulted into a network of great rivers and lakes that possess big potential for development. These resources are being developed and depleted at a fast rate, a situation that requires assessment to establish present status of water resources in the country. The paper reviews the characteristics, availability, demand and importance of present day water resources in Uganda as well as describ- ing the various issues, challenges and management of water resources of the country. -

Important Bird Areas in Uganda. Status and Trends 2008
IMPORTANT BIRD AREAS IN UGANDA Status and Trends 2008 NatureUganda The East Africa Natural History Society Important Bird Areas in Uganda Status and Trends 2008 Compiled by: Michael Opige Odull and Achilles Byaruhanga Edited by: Ambrose R. B Mugisha and Julius Arinaitwe Map illustrations by: David Mushabe Graphic designs by: Some Graphics Ltd January 2009 Reproduction of this publication for educational or other non commercial purposes is authorized without further written permission from the copyright holder provided the source is fully acknowledged. Production of this publication for resale or other commercial purposes is prohibited without prior written notice of the copyright holder. Citation: NatureUganda (2009). Important Bird Areas in Uganda, Status and Trends 2008. Copyright © NatureUganda – The East Africa Natural History Society About NatureUganda NatureUganda is a Non Governmental Organization working towards the conservation of species, sites and habitats not only for birds but other taxa too. It is the BirdLife partner in Uganda and a member of IUCN. The organization is involved in various research, conservation and advocacy work in many sites across the country. These three pillars are achieved through conservation projects, environmental education programmes and community involvement in conservation among others. All is aimed at promoting the understanding, appreciation and conservation of nature. For more information please contact: NatureUganda The East Africa Natural History Society Plot 83 Tufnell Drive, Kamwokya. P.O.Box 27034, Kampala Uganda Email [email protected] Website: www.natureuganda.org DISCLAIMER This status report has been produced with financial assistance of the European Union (EuropeAid/ ENV/2007/132-278. The contents of this document are the sole responsibility of Birdlife International and can under no normal circumstances be regarded as reflecting the position of the European Union. -

The Charcoal Grey Market in Kenya, Uganda and South Sudan (2021)
COMMODITY REPORT BLACK GOLD The charcoal grey market in Kenya, Uganda and South Sudan SIMONE HAYSOM I MICHAEL McLAGGAN JULIUS KAKA I LUCY MODI I KEN OPALA MARCH 2021 BLACK GOLD The charcoal grey market in Kenya, Uganda and South Sudan ww Simone Haysom I Michael McLaggan Julius Kaka I Lucy Modi I Ken Opala March 2021 ACKNOWLEDGEMENTS The authors would like to thank everyone who gave their time to be interviewed for this study. They would like to extend particular thanks to Dr Catherine Nabukalu, at the University of Pennsylvania, and Bryan Adkins, at UNEP, for playing an invaluable role in correcting our misperceptions and deepening our analysis. We would also like to thank Nhial Tiitmamer, at the Sudd Institute, for providing us with additional interviews and information from South Sudan at short notice. Finally, we thank Alex Goodwin for excel- lent editing. Interviews were conducted in South Sudan, Uganda and Kenya between February 2020 and November 2020. ABOUT THE AUTHORS Simone Haysom is a senior analyst at the Global Initiative Against Transnational Organized Crime (GI-TOC), with expertise in urban development, corruption and organized crime, and over a decade of experience conducting qualitative fieldwork in challenging environments. She is currently an associate of the Oceanic Humanities for the Global South research project based at the University of the Witwatersrand in Johannesburg. Ken Opala is the GI-TOC analyst for Kenya. He previously worked at Nation Media Group as deputy investigative editor and as editor-in-chief at the Nairobi Law Monthly. He has won several journalistic awards in his career. -

———— “Mudo”: the Soga 'Little Red Riding Hood'
LILLIAN BUKAAYI TIBASIIMA ———— º “Mudo”: The Soga ‘Little Red Riding Hood’ ABSTRACT This essay analyses the social underpinnings of the oral tale of “Mudo,” which belongs to the Aarne–Thompson tale type 333, along with a group of similar tales that resemble the action and movement of “Little Red Riding Hood.” Basic to the exposition is Adolf Bastian’s assertion of the fundamental similarity of ideas between all social groups. In the “Mudo” story and its Ugandan variants, the victim is a solitary little girl and the villain a male ogre who devises ways of eating her; the ogre is mostly successful, although in some variants the girl manages to escape. Although these tales come from a great range of cultures and different geographical locations, and the counterpart of the ogre in the European tales is a wolf in disguise, they share elements of plot, characteriza- tion, and motif, and address similar concerns. Introduction USOGA IS PART OF EAS TERN UGANDA, surrounded by water. The B Rev. Fredrick Kisuule Kaliisa1 notes: To the west is river Kiira (Nile) marking the boundary between Buganda and Busoga. To the East is river Mpologoma separating Busoga from Bukedi. To the North are river Mpologoma and Lake Kyoga, forming the boundary be- tween Busoga and Lango. To the south, is Lake Victoria (Nalubaale). It might be the result of the geographical location of Busoga that ogre stories were composed to warn the people against impending harm if they went out alone and stayed in secluded places. Nnalongo Lukude emphasizes this: Historically, Busoga was surrounded by bodies of water and forests, it was very bushy and as a result harboured many wild animals, some of which were man-eaters. -

Uganda and the Nile Basin Initiative: Benefits of Cooperation
UGANDA INVESTMENT BENEFITS FROM THE NILE BASIN COOPERATION ONE OF THE FOUR (4) MODERN EQUIPPED SURVEILLANCE BOATS LEAF II PROVIDED TO THE D.R CONGO AND UGANDA FOR JOINT SURVEILLANCE OF TRANSBOUNDARY LAKES EDWARD AND ALBERT LEAF II SUPPLIED TWO (2) MODERN MOBILE WATER QUALITY LABARATORY VEHICLES NEW MBARARA SUBSTATION IN UGANDA CONSTRUCTED THROUGH INTERCONNECTION TO D.R CONGO AND UGANDA TO SUPPORT WATER QUALITY MANAGEMENT OF ELECTRIC GRIDS OF NILE EQUATORIAL LAKES COUNTRIES PROJECT NBI/NELSAP–CU, APRIL 2019 MIRAMA SUBSTATION IN UGANDA CONSTRUCTED THROUGH INTERCONNECTION OF ELECTRIC GRIDS OF NILE EQUATORIAL LAKES COUNTRIES PROJECT ABOUT THE NILE EQUATORIAL LAKES SubSIDIARY ACTION PROGRAM (NELSAP) The Nile Equatorial Lakes Subsidiary Action Program regional trans-boundary projects, which are at various levels of Coordination Unit (NELSAP-CU) headquartered in Kigali, Rwanda, preparation and implementation. NELSAP-CU mobilized is one of the two investment programs of the Nile Basin Initiative USD 557.107 million cumulative finance to-date for pre- (NBI), the other being the Eastern Nile Subsidiary Action Program investment programs of and additional USD 493.018 million for (ENSAP), headquartered in Addis Ababa, Ethiopia known as investment projects. Eastern Nile Technical Regional Office (ENTRO). Since 2014, NELSAP-CU has gained regional experience, NELSAP-CU was established in December 1999 by the Council of strengthened its capacity and emerged as a reliable regional Ministers for Water Affairs with a mission to “contribute to the institution -
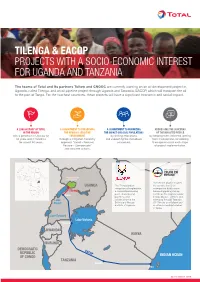
Tilenga & Eacop Projects with a Socio-Economic Interest for Uganda
TILENGA & EACOP PROJECTS WITH A SOCIO-ECONOMIC INTEREST FOR UGANDA AND TANZANIA The teams of Total and its partners Tullow and CNOOC are currently working on an oil development project in Uganda, called Tilenga, and an oil pipeline project through Uganda and Tanzania, EACOP, which will transport the oil to the port of Tanga. For the two host countries, these projects will have a significant economic and social impact. A LONG HISTORY OF TOTAL A COMMITMENT TO PRESERVING A COMMITMENT TO MINIMIZING ADDRESSING THE CONCERNS IN THE REGION THE REGION'S SENSITIVE THE IMPACT ON LOCAL POPULATIONS OF THE IMPACTED PEOPLE with a presence in Uganda for ENVIRONMENT by limiting relocations by keeping them informed, getting 60 years and in Tanzania through a mitigation hierarchy and supporting the individuals them involved and considering for almost 50 years. approach “Avoid – Reduce/ concerned. their opinions into each stage Restore – Compensate” of project implementation. and concrete actions. SOUDAN DU SUD ÉTHIOPIE Murchison Falls National Park The EACOP project involves UGANDA The Tilenga project the construction of an comprises oil exploration, underground hydrocarbon Tilenga a crude oil processing transport pipeline starting Hoima plant, underground just inside the Uganda border pipelines, and (Hoima District - 297km) and Lake infrastructure in the extending through Tanzania Albert Buliisa and Nwoya (1147km) to an oil depot and districts of Uganda. an offshore loading terminal in Tanga. Lake Edward Lake Victoria Bukoba RWANDA KENYA BURUNDI DEMOCRATIC EACOP REPUBLIC INDIAN OCEAN OF CONGO TANZANIA Singida Tanga SEPTEMBER 2019 ZAMBIE MOZAMBIQUE MALAWI SOUDAN DU SUD THIOPIE FOCUS ON THE TILENGA PROJECT Total E&P Uganda, fully aware of the project's sensitive nature, has placed particular emphasis on environmental and societal issues, with a specific commitment to leaving the site in a better state than it was before the work started and to limiting residents' relocations as much as possible. -

Impact Evaluation: Lake Victoria Transport Corridor Project
RWANDA Impact Evaluation: Lake Victoria Transport Corridor Project What is the impact of the highway construction on travel costs and aggregate economic activity? Context Lake Victoria Transport Despite recent growth, Rwanda remains amongst the poorest Program — SOP1, Rwanda countries in the world. As a landlocked country, high transport IMPLEMENTING AGENCY: costs are a critical constraint to growth. According to the Business Rwanda Transport Development Environment and Enterprise Performance (BEEP) 2011 report, about Agency (RTDA) 60 percent of firms in Rwanda rely on imports for inputs and/or supplies, which take an average of 15 days to clear customs. INTERVENTION: Construction of 119km of new, high- The Government of Rwanda (GoR) has assigned fundamental capacity highway in the Eastern and importance to the development of the economic infrastructure of Southern provinces, replacing the the country, and particularly to road transportation. The construction existing gravel road and providing of the Ngoma-Nyanza highway is identified as a priority investment new connectivity to the Tanzania port for the government under the most recent poverty reduction of entry. strategy. As depicted in blue in Figure 1, this new highway consists of two separate segments that will replace the existing gravel road PROJECT DEVELOPMENT OBJECTIVE and will provide new connectivity within the National Road System To contribute to the efficient and between the Tanzania port of entry in the south east of the country safe movement of goods and people and southern central Rwanda. The first section has a length of along the regional corridor from the border crossing at Rusumo to the border crossing at Nemba. -

The Precolonial Social Formation Among the Bakenhe Fishing Community
The Precolonial Social Formation Among the Bakenhe Fishing Community .. of Lake Kyoga Il.egion of Uganda , 1800- 1894* • By C. Asowa- Okwe , Department of Political Science and Public Adminis t rat i on. Int roduction It i s eminently evident from the literqture ava ilable on the pre- colonia l history of Uganda and East Africa in general , that one area which has either been negl ected , or peripher ally treat ed i s that of the fishing industry and the fishing communities . A car eful and close examina tion of these liter a ture show a definite bia s towar ds the agricult ural and pastoral communities and their economic activities. And as if tha t is not enough , f ew that have t r i ed to grappl e with the fishing industry have l ar gely tended to be descriptive/narra tive , and ma inly, t alking a bout methods of fishing and the types of fis hes c aught. In f act the bulk of literature ~ on the fishing industry are basically , works of the physical scientists who ar e mainl y tra i ned in bi ol ogica l sciences . These studies , ther ef or e , ma i nly focus on the fish species f ound in the wa t ers of East African l akes , rivers , ponds and swamps , their f ood r equirements and dist ribut ion. They a lso concern thems elves with the question of density of fish popu- l a tion, the growth r a t e of i ndividua l species, the age at which they mature , t he specific f actor s which cause ornt~o p ic a l l ake to support many fish and another r el a tively f ew, the depletion of certa in fish species , the stocking of new species, and how t o check .the depl etion of s ome species, like·, ales.tes ( soga) , Labeo (ningu), bagrus (semutundu) . -

Characteristics of Macrophytes in the Lubigi Wetland in Uganda
Vol. 10(10), pp. 394-406, October 2018 DOI: 10.5897/IJBC2018.1206 Article Number: 98C504658827 ISSN: 2141-243X Copyright ©2018 International Journal of Biodiversity and Author(s) retain the copyright of this article http://www.academicjournals.org/IJBC Conservation Full Length Research Paper Characteristics of macrophytes in the Lubigi Wetland in Uganda John K. Kayima and Aloyce W. Mayo* Department of Water Resources Engineering, University of Dar es Salaam, P. O. Box 35131, Dar es Salaam, Tanzania. Received 1 July, 2018; Accepted 16 August, 2018 The Lubigi wetland, which is located in the north-western part of Kampala, the capital city of Uganda has been severely strained from anthropogenic encroachment and activities. These activities include harvesting of Cyperus papyrus and other plants, land filling for reclamation, human settlements and disposal of wastewater into the wetland among others. As a result of these anthropogenic activities, the macrophytes diversity and biomass in the wetland have been affected, which in turn affects the effectiveness of wetland for removal of pollutants. It is therefore important to investigate the characteristics of wetland macrophytes in the Lubigi wetland. Pertinent field investigations, surveys, data collection and laboratory tests and analyses were carried out. The problem being addressed was the current lack of information and knowledge about the biomass and biodiversity of the Lubigi wetland to protect the downstream Mayanja River and Lake Kyoga. Three transects each of 1.0 m wide was cut across this zone at about 700 m downstream of the main wastewater inlet, the second at about 1,440 m downstream of the main wastewater inlet and the third at about 1,930 m downstream of the main wastewater inlet.