Impacts of Ocean Climate Variability on Biodiversity of Pelagic Forage Species in an Upwelling Ecosystem
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Forage Fish Management Plan
Oregon Forage Fish Management Plan November 19, 2016 Oregon Department of Fish and Wildlife Marine Resources Program 2040 SE Marine Science Drive Newport, OR 97365 (541) 867-4741 http://www.dfw.state.or.us/MRP/ Oregon Department of Fish & Wildlife 1 Table of Contents Executive Summary ....................................................................................................................................... 4 Introduction .................................................................................................................................................. 6 Purpose and Need ..................................................................................................................................... 6 Federal action to protect Forage Fish (2016)............................................................................................ 7 The Oregon Marine Fisheries Management Plan Framework .................................................................. 7 Relationship to Other State Policies ......................................................................................................... 7 Public Process Developing this Plan .......................................................................................................... 8 How this Document is Organized .............................................................................................................. 8 A. Resource Analysis .................................................................................................................................... -

Crestfish Lophotus Lacepede (Giorna, 1809) and Scalloped Ribbonfish Zu Cristatus (Bonelli, 1819) in the Northern Coast of Sicily, Italy
ISSN: 0001-5113 ACTA ADRIAT., ORIGINAL SCIENTIFIC PAPER AADRAY 58(1): 137 - 146, 2017 Occurrence of two rare species from order Lampriformes: Crestfish Lophotus lacepede (Giorna, 1809) and scalloped ribbonfish Zu cristatus (Bonelli, 1819) in the northern coast of Sicily, Italy Fabio FALSONE1, Michele Luca GERACI1, Danilo SCANNELLA1, Charles Odilichukwu R. OKPALA1, Giovan Battista GIUSTO1, Mar BOSCH-BELMAR2, Salvatore GANCITANO1 and Gioacchino BONO1 1Institute for the Coastal Marine Environment, IAMC‑CNR, 91026 Mazara del Vallo, Sicily, Italy 2Consorzio Nazionale Interuniversitario per le Scienze del Mare (CoNISMa), Rome, Italy Corresponding author, e‑mail: [email protected] The bony fish Lophotus lacepede (Giorna, 1809) and Zu cristatus (Bonelli, 1819) are the two species rarely recorded within the Mediterranean basin, usually reported as accidentally captured in depth (mesopelagic) fishing operations. In the current work, we present the first record of L. lacepede and Z. cristatus in fishing catches from southwestern Tyrrhenian Sea. Moreover, in order to improve existent biological/ecological knowledge, some bio-related aspects such as feeding aspect, sexual maturity and age estimate have been discussed. Key words: crestfish, scalloped ribbonfish, meristic features, vertebrae, growth ring INTRODUCTION species of Lophotidae family, the L. lacepede inhabits the epipelagic zone, although it could The target species of this study (Lophotus also be recorded in most oceans from the surface lacepede and Zu cristatus) belong to Lophotidae up to 1000 m depth (HEEMSTRA, 1986; PALMER, (Bonaparte, 1845) and Trachipteridae (Swain- 1986; OLNEY, 1999). First record of this spe- son, 1839) families respectively, including the cies in the Mediterranean Basin was from the Lampriformes order (consisted of 7 families). -

Fishery Bulletin/U S Dept of Commerce National Oceanic
Abstract.- Gastrointestinal tract contents were evaluated from Prey selection by northern fur seals 73 female andjuvenile male north ern fur seals (Callorhinus ursinus) fCallorhinus ursinusJ in the eastern for analysis of their diet in the Bering Sea. Fur seals were col Bering Sea lected from August to October of 1981, 1982, and 1985. Juvenile Elizabeth Sinclair walleye pollock (Theragra chalco gramma) and gonatid squid were Thomas Loughlin the primary prey. Pacific herring National Marine Mammal Laboratory. Alaska Fisheries Science Center; (Clupea pallasi) and capelin National Marine Fisheries Service. NOM <Mallotus villosus), considered im 7600 Sand Point Way. N.E., Seattle, Washington 98115 portant fur seal prey in previous reports, were absent from the diet. Prey species and size varied William Pearcy among years and between near College of Oceanography, Oregon State University shore and pelagic sample loca Oceanography Administration Building 104 tions. Interannual variation in the Corvaflis, Oregon 9733 J importance ofpollock in the diet of fur seals was positively related to year-class strength of pollock. Midwater (n=23) and bottom (n=116) trawls were conducted at The Pribilof Island population (St. were conducted prior to the 1975 the location of fur seal collections George and St. Paul Islands) of 81 population decline and prior to to determine availability of fish northern fur seals (Callorhinus the 1970's development of a com and squid relative to prey species eaten by fur seals. The species and ursinus) represents approximately mercial walleye pollock (Theragra size composition of prey taken by 75% of the total species breeding chalcogramma) fishery in the fur seals was similar to midwater population. -

Full Text in Pdf Format
MARINE ECOLOGY PROGRESS SERIES Vol. 177: 115-131, 1999 Published February 11 Mar Ecol Prog Ser Diet, provisioning and productivity responses of marine predators to differences in availability of Antarctic krill J. P. Croxall*, K. Reid, P. A. Prince British Antarctic Survey, Natural Environment Research Council. High Cross, Madingley Road. Cambridge CB3 OET. United Kingdom ABSTRACT: Knowledge of relationships between prey availability and predator performance is the key to using predators as indicators of the state of marine systems and to assessing potential conse- quences of competition between natural predators and man for common resources. Fluctuations in the abundance of Antarctic knll are beiieved to have a substantial influence on the reproductive perfor- mance of krill-dependent top predator species in the Southern Ocean; few quantifications of such inter- actions exist. At South Georgia, for 2 years in which acoustic surveys revealed a major difference in knll abundance, we compared diet, prov~sioningof offspring and breeding success in 4 main predator species (2 penguins, 2 albatrosses, with supporting data from Antarctic fur seal) whose dependence on krd typically ranges from 20 to 90%. The 4-fold difference in krill biomass between 1986 (ca 30 g m-2) and 1994 (ca 7 g m-2)was accompanied by (1) an 88 to 90% reduction in the mass of krill in predator dlets (and some increase in the fish component), (2) greater prey diversity for most species, (3) reduced diet overlap between species and (4) a switch from krill to amphipods in macaroni penguin but no major dietary change in other species. -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

Long-Term Fluctuations in Gonatid Squid (Gonatidae) Abundance in the Okhotsk Sea
Pacific Research Institute of Fisheries and Oceanography (TINRO-Centre) Long-term fluctuations in gonatid squid (Gonatidae) abundance in the Okhotsk Sea Mikhail A. Zuev Nikolai S. Vanin Oleg N. Katugin Gennady A. Shevtsov The main goal reveal annual distribution patterns of squids of the family Gonatidae in the Sea of Okhotsk basing on data collected in TINRO-Centre research surveys with a purpose to estimate the role of these cephalopods in pelagic communities Methods: – analyze database on squid catches for a period from 1990 to 2008; – use the data to obtain patterns of spatial and seasonal on squid distribution and biology; – assess the role of different species in seasonal dynamics of distribution density in the epi- and mesopelagic layers in different years; – use synoptic observations to reveal relationships between squid distribution density and variability in atmospheric pressure in the study area Upper-mesopelagic Region expeditions investigations years (expeditions) northern Okhotsk Sea 20 12 1990-2008 southern 18 2 1990-2008 Okhotsk Sea off west 12 7 1998-2007 Kamchatka the entire Okhotsk Sea 25 13 1990-2008 - Investigations of epipelagic (0-200 m) - Investigations of epipelagic and upper- mesopelagic (200-500 m) northern Okhotsk Sea Kamchatka Biomass density (kg/km2) of the Gonatidae in the northern Okhotsk Sea kg/km2 0,6 2 - Berryteuthis magister 0,5 4000,4 - Gonatus madokai 0,3 2000,2 0,1 0 w w s s о w s s о w s s о w s s о w s s о о w s s о w s s s о 1991 ] [ 1993][1994 1995 ][ 1997 1998 1999 2000 1,2 - Boreoteuthis borealis -

An Isotope Approach
Determining Cetacean – Cephalopod Trophic Interactions – An Isotope Approach By Christine Heather Jackson Bachelor of Arts, James Cook University Graduate Diploma (Health Promotion), Curtin University of Technology Master of Science, University of Tasmania Submitted in fulfilment of the requirements for the degree Doctor of Philosophy University of Tasmania (October 2016) ii DECLARATION This thesis contains no material which has been accepted for a degree or diploma by the University or any other institution, except by way of background information and duly acknowledged in the thesis, and to the best of my knowledge and belief no material previously published or written by another person except where due acknowledgement is made in the text of the thesis, nor does the thesis contain any material that infringes copyright Signed; _________________________________ Christine H. Jackson iii AUTHORITY OF ACCESS This thesis may be made available for loan and limited copying and communication in accordance with the Copyright Act 1968. Signed; _________________________________ Christine H. Jackson iv ABSTRACT Determining the biotic and abiotic influences on the distribution and abundance of marine mammals is essential for understanding the dynamics of the food chain. The predator-prey relationship can be deterministic in shaping both the community structure and function of marine ecosystems. This is especially pertinent to recovering toothed whale populations given their large size and high prey consumption rates. A greater knowledge of the trophic linkages between toothed whales and their prey will facilitate assessments of their combined impact on the ecosystem since marine food webs are a fusion of bottom-up and top-down energy and nutrient flow. -

Redacted for Privacy William G
AN ABSTRACT OF THE THESIS OF Elizabeth H. Sinclair for the degree of Master of Science in Oceanography presented on December 16, 1988. Title: Feeding Habits of Northern Fur Seals (Callorhinus ursinus) in the Eastern Bering Sea Abstract approved: Redacted for privacy William G. Pearcy This study was conducted to determine the composition and size of prey consumed by northern fur seals (Callorhinus ursinus) in the eastern Bering Sea. Eighty three northern fur seals were collected in the summer and fall of 1981, 1982, and 1985 forexamination of gastrointestinal contents. A total of 139 midwater and bottom trawls were collected to determine the availability of potential prey. Analysis of trawls confirmed that seals are size-selective midwater feeders during their breeding and haul-out season in the eastern Bering Sea. Juvenile walleye pollock and gonatid squid, 5-20cm in body length, were the primary prey, but seal prey varied among years and between nearshore and pelagic sample locations. Interannual variation in body sizes of walleye pollock consumed by seals was related to pollock year class strength. The identification of pollock and gonatid squid as primary fur seal prey in the eastern Bering Sea was consistent with previous reports. However, Pacific herring and capelin, previously considered important fur seal prey werE absent in this study. Feeding Habits of Northern Fur Seals (Callorhinus ursinus) in the Eastern Bering Sea by Elizabeth Hacker Sinclair A THESIS submitted to Oregon State University in partial fulfillment of the requirements for the degree of Master of Science Completed December 16, 1988 Commencement June 1989 APPROVED: Redacted for privacy Professor of Oceanography in char.4f major Redacted for privacy Dean of C./lege of Ocean. -

Fish Bulletin 161. California Marine Fish Landings for 1972 and Designated Common Names of Certain Marine Organisms of California
UC San Diego Fish Bulletin Title Fish Bulletin 161. California Marine Fish Landings For 1972 and Designated Common Names of Certain Marine Organisms of California Permalink https://escholarship.org/uc/item/93g734v0 Authors Pinkas, Leo Gates, Doyle E Frey, Herbert W Publication Date 1974 eScholarship.org Powered by the California Digital Library University of California STATE OF CALIFORNIA THE RESOURCES AGENCY OF CALIFORNIA DEPARTMENT OF FISH AND GAME FISH BULLETIN 161 California Marine Fish Landings For 1972 and Designated Common Names of Certain Marine Organisms of California By Leo Pinkas Marine Resources Region and By Doyle E. Gates and Herbert W. Frey > Marine Resources Region 1974 1 Figure 1. Geographical areas used to summarize California Fisheries statistics. 2 3 1. CALIFORNIA MARINE FISH LANDINGS FOR 1972 LEO PINKAS Marine Resources Region 1.1. INTRODUCTION The protection, propagation, and wise utilization of California's living marine resources (established as common property by statute, Section 1600, Fish and Game Code) is dependent upon the welding of biological, environment- al, economic, and sociological factors. Fundamental to each of these factors, as well as the entire management pro- cess, are harvest records. The California Department of Fish and Game began gathering commercial fisheries land- ing data in 1916. Commercial fish catches were first published in 1929 for the years 1926 and 1927. This report, the 32nd in the landing series, is for the calendar year 1972. It summarizes commercial fishing activities in marine as well as fresh waters and includes the catches of the sportfishing partyboat fleet. Preliminary landing data are published annually in the circular series which also enumerates certain fishery products produced from the catch. -

First Record of Polka-Dot Ribbonfish from India 3
Marine Biodiversity Records, page 1 of 4. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S1755267211001151; Vol. 5; e8; 2012 Published online First record of Polka-dot ribbonfish Desmodema polystictum (Pisces: Trachipteridae) from Indian waters p.u. zacharia and k. kannan Central Marine Fisheries Research Institute, PB No.1603, Ernakulam North PO, Cochin-682 018, Kerala, India Polka-dot ribbonfish Desmodema polystictum was recorded for the first time from Indian waters. A single specimen of D. polystictum (107 cm total length and weighing 480 g) was collected from Tharuvaikulam landing centre, north to Tuticorin, on the south-east coast of India during September 2010. The distinguishing characters of the species from other species of the family are discussed. Morphometric and meristic characters of D. polystictum are presented in this paper. With the present report, the distribution area of this species now extends to the Indian waters. Keywords: first record, Polka-dot ribbonfish, Desmodema polystictum, Indian waters Submitted 6 September 2011; accepted 29 November 2011 INTRODUCTION of D. polystictum aresilverywithprofusedarkspotting(polka dotted) but the adults lack spots. The fish of the family Trachipteridae are characterized by long Froese & Pauly (2010) state that Desmodema polystictum compressed ribbon or tape-shaped body, short head, and probably has a circumtropical distribution; it was reported narrow mouth (Heemstra & Kannemeyer, 1986). The pectoral from Japan, Taiwan (Shen, 1993), Philippines, Australia, fin is small, pelvic long and fan like in young composed of New Zealand (Paulin et al., 1989); Western Pacific (Ogilby, several rays, absent in adults. Anal fin absent. These fish also 1897) and 16811′N to Namibia (Aguiar & Que´ro, 1990); have a high dorsal fin that actually occupies the entire length South Africa in the eastern Atlantic; Florida, USA (Moore of its back with origin well behind tip of snout. -
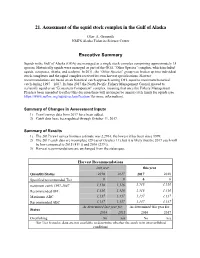
Assessment of the Squid Stock Complex in the Gulf of Alaska
21. Assessment of the squid stock complex in the Gulf of Alaska Olav A. Ormseth NMFS Alaska Fisheries Science Center Executive Summary Squids in the Gulf of Alaska (GOA) are managed as a single stock complex comprising approximately 15 species. Historically squids were managed as part of the GOA “Other Species” complex, which included squids, octopuses, sharks, and sculpins. In 2011, the “Other Species” group was broken up into individual stock complexes and the squid complex received its own harvest specifications. Harvest recommendations are based on an historical catch approach setting OFL equal to maximum historical catch during 1997 – 2007. In June 2017 the North Pacific Fishery Management Council moved to reclassify squid as an “Ecosystem Component” complex, meaning that once the Fishery Management Plan has been amended to reflect this decision there will no longer be annual catch limits for squids (see https://www.npfmc.org/squid-reclassification/ for more information). Summary of Changes in Assessment Inputs 1) Trawl survey data from 2017 have been added. 2) Catch data have been updated through October 11, 2017. Summary of Results 1) The 2017 trawl survey biomass estimate was 2,296 t, the lowest it has been since 1999. 2) The 2017 catch data are incomplete (29 t as of October 11), but it is likely that the 2017 catch will be low compared to 2015 (411 t) and 2016 (239 t). 3) Harvest recommendations are unchanged from the status quo. Harvest Recommendations last year this year Quantity/Status 2016 2017 2017 2018 Specified/recommended -

Training Manual Series No.15/2018
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by CMFRI Digital Repository DBTR-H D Indian Council of Agricultural Research Ministry of Science and Technology Central Marine Fisheries Research Institute Department of Biotechnology CMFRI Training Manual Series No.15/2018 Training Manual In the frame work of the project: DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals 2015-18 Training Manual In the frame work of the project: DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals 2015-18 Training Manual This is a limited edition of the CMFRI Training Manual provided to participants of the “DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals” organized by the Marine Biotechnology Division of Central Marine Fisheries Research Institute (CMFRI), from 2nd February 2015 - 31st March 2018. Principal Investigator Dr. P. Vijayagopal Compiled & Edited by Dr. P. Vijayagopal Dr. Reynold Peter Assisted by Aditya Prabhakar Swetha Dhamodharan P V ISBN 978-93-82263-24-1 CMFRI Training Manual Series No.15/2018 Published by Dr A Gopalakrishnan Director, Central Marine Fisheries Research Institute (ICAR-CMFRI) Central Marine Fisheries Research Institute PB.No:1603, Ernakulam North P.O, Kochi-682018, India. 2 Foreword Central Marine Fisheries Research Institute (CMFRI), Kochi along with CIFE, Mumbai and CIFA, Bhubaneswar within the Indian Council of Agricultural Research (ICAR) and Department of Biotechnology of Government of India organized a series of training programs entitled “DBT sponsored Three Months National Training in Molecular Biology and Biotechnology for Fisheries Professionals”.