River Flow Impacts on Estuarine Prawns in the Gulf of Carpentaria
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Improved Continuing Losses Estimation Using Initial Loss-Continuing Loss Model for Medium Sized Rural Catchments
American J. of Engineering and Applied Sciences 2 (4): 796-803, 2009 ISSN 1941-7020 © 2009 Science Publications Improved Continuing Losses Estimation Using Initial Loss-Continuing Loss Model for Medium Sized Rural Catchments Mahbub Ilahee and Monzur Alam Imteaz Faculty of Engineering and Industrial Sciences, Swinburne University of Technology, Hawthorn, Melbourne, VIC 3122, Australia Abstract: Problem statement: The rainfall based design flood estimation techniques are commonly adopted in hydrological design and require a number of inputs including information on soil loss characteristics. Approach: A conceptual loss model known as the ‘Initial Loss-Continuing Loss (IL- CL) model’ is widely used in Australia. Results: The Initial Loss (IL) occurs at the beginning of the rainfall event, prior to the commencement of surface runoff and the Continuing Loss (CL) is the average rate of loss throughout the remainder of the storm. The currently recommended design loss values depicted in “Australian Rainfall and Runoff Vol. 1” for Queensland (Australia) has some basic limitations. This study investigated how more accurate CL values can be estimated and derived for medium sized tropical Queensland catchments using long term rainfall and streamflow data. Accuracy in CL estimation has got significant implications in the estimation of design floods. Conclusion/Recommendations: The results showed that CL value is not fixed and constant through out the duration of the storm but the CL value decays with the duration of the storm. Key words: Initial loss, continuing loss, rural catchments, flood estimation, rainfall-runoff modeling PROBLEM STATEMENT topography, soil characteristics, vegetation and climate; the components exhibit a high degree of temporal and Flood estimation is often required in hydrologic spatial variability during high rainfall events. -

Surface Water Ambient Network (Water Quality) 2020-21
Surface Water Ambient Network (Water Quality) 2020-21 July 2020 This publication has been compiled by Natural Resources Divisional Support, Department of Natural Resources, Mines and Energy. © State of Queensland, 2020 The Queensland Government supports and encourages the dissemination and exchange of its information. The copyright in this publication is licensed under a Creative Commons Attribution 4.0 International (CC BY 4.0) licence. Under this licence you are free, without having to seek our permission, to use this publication in accordance with the licence terms. You must keep intact the copyright notice and attribute the State of Queensland as the source of the publication. Note: Some content in this publication may have different licence terms as indicated. For more information on this licence, visit https://creativecommons.org/licenses/by/4.0/. The information contained herein is subject to change without notice. The Queensland Government shall not be liable for technical or other errors or omissions contained herein. The reader/user accepts all risks and responsibility for losses, damages, costs and other consequences resulting directly or indirectly from using this information. Summary This document lists the stream gauging stations which make up the Department of Natural Resources, Mines and Energy (DNRME) surface water quality monitoring network. Data collected under this network are published on DNRME’s Water Monitoring Information Data Portal. The water quality data collected includes both logged time-series and manual water samples taken for later laboratory analysis. Other data types are also collected at stream gauging stations, including rainfall and stream height. Further information is available on the Water Monitoring Information Data Portal under each station listing. -

The Freshwater Crayfish (Family Parastacidae) of Queensland
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Riek, E. F., 1951. The freshwater crayfish (family Parastacidae) of Queensland. Records of the Australian Museum 22(4): 368–388. [30 June 1951]. doi:10.3853/j.0067-1975.22.1951.615 ISSN 0067-1975 Published by the Australian Museum, Sydney nature culture discover Australian Museum science is freely accessible online at http://publications.australianmuseum.net.au 6 College Street, Sydney NSW 2010, Australia 11ft! FRESHWATER CRAYFISH (FAMILY PARASTACIDAE) OF QUEENSLAND WITH AN ApPENDIX DESORIBING OTHlm AV5'lHALIAN SPEClEf'. By E. F. HIEK. (;ommonwealth Scientific and Industrial l~csearch Organization - Divhdon of Entomology, Canberra, A.C.T. (Figures 1-13.) Freshwater crayfish occur in almost every body of fresh water from artificial damfl and natural billabongs (I>tanding water) to headwater creeks and large rivers (flowing water). Generally the species are of considerable size and therefore easily collected, but even so many of the larger forms are unknown scientifically. This paper deals with all the species that have been collected from Queensland. It also includes a few species from New South Wales and other States. No doubt additional species will be found and some of the mOre variable series, at present included under the one specific namc, will be further subdivided. From Queensland nine species are described as new, making a total of seventeen species (of three genera) recorded from that State. The type localities of all but two of these species are in Queensland but some are not restricted to the State. Clark's 1936 and subsequent papers have been used as the basis for further taxonomic studies of the Australian freshwater crayfish. -

Port of Karumba, July 2021
4. Port description 4.1 General Information Karumba is situated in the south-east corner of the Gulf of Carpentaria, 530 kilometres west of Cairns at the mouth of the Norman River. The Karumba Shipping Channel has been maintained in the past for Zinc concentrate exports. The mine was closed in February 2016 and there will be no more large scale maintenance dredging of the shipping channel. Far North Queensland Ports Corporation Limited, who manage the port, have advised that they will not be commissioning maintenance dredging therefore it is expected that the channel will gradually reduce in depth and width due to siltation. A small scale maintenance dredging campaign took place in 2018 to remove siltation for the commencement of limited zinc concentrate exports. Studies undertaken by Far North Queensland Ports Corporation Limited indicate that the minimum depth of the channel could be reduced to -2.0m LAT within 2 years – the rate of siltation may be effected by the strength of the NW monsoon during the wet season. Karumba also provides a trawler base for the prawn and fishing industry, the export of live cattle and a community port for servicing townships in the area. 4.2 Pilotage area The Karumba Pilotage area is described in Schedule 2 of the Transport Operations (Marine Safety) Regulation 2016 as the area of: a) Waters at the high water mark consisting of the following: • the Norman River and connected waterways system from the head of navigation to the river mouth • from the river mouth, the waters within a 10 nautical mile radius centred at the north head of the Norman River entrance; and The navigable waters of rivers and creeks flowing, directly or indirectly, into the waters in paragraph (a). -

The Gulf Savannah Is a Far Medical Centres at Georgetown, Forsayth, Normally in Force from October to February
Head Office: Department of Natural Resources and Water Cnr Main & Vulture Sts, Woolloongabba, Brisbane Locked Bag 40, Coorparoo Delivery Centre, Qld. 4151 Ph (07) 3896 3216, Fax (07) 3896 3510 For all your regional and recreational map needs, Sunmap products are available from Departmental service centres, distributors and selected retailers throughout Queensland or the Queensland Government Bookshop at: www.publications.qld.gov.au. The development of aviation and the inspiration of John Flynn To view the complete range of products and services, visit our home combined after World War I to include the remote Gulf frontier in page at: www.nrw.qld.gov.au. the network of Flying Doctor Services which made up the ‘mantle of safety’ for the inland areas of Australia. The Etheridge Goldfield, the ‘poor man’s goldfield’ has never been worked out. Discovered by Richard Daintree in 1869, the Etheridge survived the rushes to the Palmer Over the bush ‘roads’ rolled the legendary and other richer fields in North Queensland. The ghosts Founded in 1865 by commercial and pastoral interests led by The traditional industries of the Gulf The Normanton to Croydon Railway is a living relic of the age of steam railways. Originally coaches of Cobb and Co. and other lines, of such towns as Charleston on the Etheridge and Robert Towns, Burketown in its early days was a wild frontier Savannah are fishing and grazing, with intended to link the port of Normanton to the copper mines of Cloncurry, the discovery of gold carrying mail and passengers between Gilberton on the Gilbert Field still dot the Savannah and Weipa town, the refuge of law breakers and adventurers, a town which beef cattle succeeding sheep, which were around Croydon led to its diversion to that Goldfield in 1891. -

Surface Water Network Review Final Report
Surface Water Network Review Final Report 16 July 2018 This publication has been compiled by Operations Support - Water, Department of Natural Resources, Mines and Energy. © State of Queensland, 2018 The Queensland Government supports and encourages the dissemination and exchange of its information. The copyright in this publication is licensed under a Creative Commons Attribution 4.0 International (CC BY 4.0) licence. Under this licence you are free, without having to seek our permission, to use this publication in accordance with the licence terms. You must keep intact the copyright notice and attribute the State of Queensland as the source of the publication. Note: Some content in this publication may have different licence terms as indicated. For more information on this licence, visit https://creativecommons.org/licenses/by/4.0/. The information contained herein is subject to change without notice. The Queensland Government shall not be liable for technical or other errors or omissions contained herein. The reader/user accepts all risks and responsibility for losses, damages, costs and other consequences resulting directly or indirectly from using this information. Interpreter statement: The Queensland Government is committed to providing accessible services to Queenslanders from all culturally and linguistically diverse backgrounds. If you have difficulty in understanding this document, you can contact us within Australia on 13QGOV (13 74 68) and we will arrange an interpreter to effectively communicate the report to you. Surface -

Three Rivers Irrigation Project Initial Advice Statement
Three Rivers Irrigation Project Initial Advice Statement June 2015 TRIP Initial Advice Statement: Stanbroke TRIP Initial Advice Statement: Stanbroke TABLE OF CONTENTS GLOSSARY ..................................................................................................................................... I EXECUTIVE SUMMARY ................................................................................................................. III 1. INTRODUCTION ...................................................................................................................... 1 1.1. Background ....................................................................................................................... 1 1.1.1. Purpose and Scope of the Initial Advice Statement ................................................. 1 2. THE PROPONENT.................................................................................................................... 3 2.1. Stanbroke Pty Ltd .............................................................................................................. 3 3. THE NATURE OF THE PROPOSAL ............................................................................................. 4 3.1. Scope of the Project .......................................................................................................... 4 3.1.1. Water Extraction ....................................................................................................... 4 3.1.2. Offstream Storages .................................................................................................. -
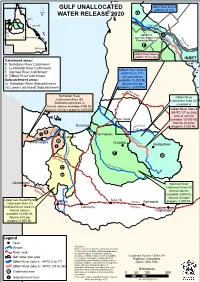
Gulf Unallocated Water Release 2020
GULF UNALLOCATED Gilbert River Zone 6 (AMTD 0 to 171) ´ WATER RELEASE 2020 G Cairns Townsville GilbertRiv er Mt Isa AMTD 171 Rockhampton (junction Gilbert and Ei Ri n Einasleigh Rivers) a v slei er F gh Brisbane Gilbert River Zone 6 (AMTD 171 to 368) INSET Catchment areas: B. Nicholson River Catchment C. Leichhardt River Catchment Gilbert River Zone 6 F. Norman River Catchment (AMTD 0 to 171) G. Gilbert River Catchment Annual volume Subcatchment areas: available 75,000 ML (i) Nicholson River Subcatchment Volume limit per (iv) Lower Leichhardt Subcatchment property 25,000 ML D e ev Burk Ro ad Nicholson River Gilbert River Catchment Area (B) Catchment Area (G) Subcatchment Area (i) unzoned or Annual volume available 4400 ML Volume limit per property 4400 ML Gilbert River Zone 6 (AMTD 171 to 368) Annual volume See Inset available 10,000 ML Volume limit per sleigh on Karumba Eina property 2,000 ML ls r o e River h iv B R i Nic Normanton Gi Burketown l bert B R Doomadgee Croydon iv er Georgetown i Le i c h ha W r F ills d t G DevRoadR ive r N orman R C iver iv Camooweal Norman River B ark ly Catchment Area (F) H ighwa Annual volume y available 3,000 ML Volume limit per Lower Leichhardt River Julia Ck. Mt Isa Flind property 3,000 ML Catchment Area (C) ersRichmond Subcatchment Area (iv) Cloncurry Highway Annual volume Hughenden available 10,000 ML Volume limit per property 5,000 ML Legend Town Disclaimer: Stream While every care is taken to ensure the accuracy of this product, the State of Queensland makes Major road no representations or warranties -

Addressing Knowledge Gaps for Studies of the Effect of Water
Addressing knowledge gaps for studies of the effect of water resource development on the future of the NPF: juvenile banana prawn abundance in estuarine habitats in the Mitchell, Gilbert and Flinders Rivers Rob Kenyon, Michele Burford, Annie Jarrett, Stephen Faggotter, Chris Moeseneder, Rik Buckworth Project No. 2016/047 i 2016/047 Addressing knowledge gaps for studies of the effect of water resource development on the future of the Northern Prawn Fishery PRINCIPAL INVESTIGATOR: Mr R. Kenyon ADDRESS: CSIRO Oceans and Atmosphere Queensland Biosciences Precinct 306 Carmody Road St Lucia QLD 4067 Telephone: 07 3833 5934 OBJECTIVES: 1. Synthesise historical data available on surveys of the fishery and recruitment of prawns. 2. Contribute to the sampling design for field trips to the southern Gulf of Carpentaria to estimate juvenile prawn densities across estuarine nursery habitats. 3. Undertake field sampling across estuaries in the southern Gulf of Carpentaria to estimate juvenile prawn densities and explore linkages to primary production in rivers with different characteristics and catchment features compared to the Norman River, expanding knowledge to other GOC rivers. 4. Contribute to data analysis from field sampling effort (Objective 2) and provide advice on sample sorting and analysis. ii NON TECHNICAL SUMMARY: OUTCOMES ACHIEVED TO DATE Accounting for past and current estuarine and offshore banana prawn survey outcomes, contributed to the sampling design for prawns, meiofauna and productivity in the Mitchell, Gilbert and Flinders Rivers during field trips from 2016 to 2019. Meiofauna and productivity sampling was undertaken by Dr. Michael Venarsky and Professor Michele Burford, colleagues (and a co-investigator) from Griffith University as part of NESP 1.4 (Links between Gulf Rivers and Coastal Productivity). -

Inland Waters Regional NRM Assessment | 2015 1
Inland Waters Regional NRM Assessment | 2015 1 Prepared by: NRM Planning @ Northern Gulf Resource Management Group Ltd Lead author: Jim Tait, Econcern Consulting Contributors: Sarah Rizvi & Natalie Waller Reviewers & Advisors: Alf Hogan, Amanda Stone, Andrew Brooks, Brynn Matthews, Damien Burrows, Jon Brodie, Jeff Shellberg, Malcolm Pearce, Mark Kennard, Michaelie Pollard, Michele Burford, Rob Ryan, Sarah Connor, Stephen Mackay, Terry Valance, Timothy Jardine Design work: Clare Powell & Federico Vanni Editing: Nina Bailey Photography: Federico Vanni This project is supported by the Northern Gulf Resource Management Group Ltd through funding from the Australian Government. Inland Waters Regional NRM Assessment | 2015 2 CONTENTS 1.1. INTRODUCTION - NORTHERN GULF FRESHWATER ECOSYSTEM NRM ISSUES ............................................ 5 1.2. ASSETS AND STATUS .................................................................................................................................................. 8 1.2.1. Freshwater Environments .................................................................................................................................... 8 1.2.2. Freshwater Biodiversity .................................................................................................................................. 38 1.2.3. Freshwater Fishery Resources ........................................................................................................................ 51 1.2.4. Water Resources ............................................................................................................................................. -

Early North Queensland
EARLY DAYS IN NORTH QUEENSLAND EARLY DAYS IN NORTH QUEENSLAND BY THE LATE EDWARD PALMER SYDNEY ANGUS & ROBERTSON MELBOURNE: ANGUS, ROBERTSON & SHENSTONE 1903 This is a blank page TO THE NORTH-WEST. I know the land of the far, fa y away, Where the salt bush glistens in silver-grey ; Where the emit stalks with her striped brood, Searching the plains for her daily food. I know the land of the far, far west, Where the bower-bird builds her playhouse nest ; Where the dusky savage from day to day, Hunts with his tribe in their old wild way. 'Tis a land of vastness and solitude deep, Where the dry hot winds their revels keep ; The land of mirage that cheats the eye, The land of cloudless and burning sky. 'Tis a land of drought and pastures grey, Where flock-pigeons rise in vast ark ay ; Where the " nardoo" spreads its silvery sheen Over the plains where the floods have beeh. 'Tis a land of gidya and dark boree, Extended o'er plains like an inland sea, Boundless and vast, where the wild winds pass, O'er the long rollers and billows of grass. I made my home in that thirsty land, Where rivers for water are filled with sand ; Where glare and heat and storms sweep by, Where the prairie rolls to the western sky. Cloncurry, 1897. —" Loranthus." W. C. Penfold & Co., Printers, Sydney. PREFACE. HE writer came to Queensland two years before T separation, and shortly afterwards took part in the work of outside settlement, or pioneering, looking for new country to settle on with stock. -

Environmental Flows for Sub-Tropical Estuaries: Understanding the Freshwater Needs for Sustainable Fisheries Production and Assessing the Impacts of Water Regulation
Queensland the Smart State Environmental flows for sub-tropical estuaries: understanding the freshwater needs of estuaries for sustainable fisheries production and assessing the impacts of water regulation Compiled by Ian Halliday and Julie Robins Final Report FRDC Project No. 2001/022 Coastal Zone Project FH3/AF C 0042 PR07-2901 ISBN 978 0 7345 0364 0 The Department of Primary Industries and Fisheries (DPI&F) seeks to maximise the economic potential of Queensland’s primary industries on a sustainable basis. This publication has been compiled by Ian Halliday and Julie Robins [Sustainable Fisheries Unit, Animal Science, Delivery]. While every care has been taken in preparing this publication, the State of Queensland accepts no responsibility for decisions or actions taken as a result of any data, information, statement or advice, expressed or implied, contained in this report. © The State of Queensland, Department of Primary Industries and Fisheries, the Coastal Zone Cooperative Research Centre and the Fisheries Research Devlopment Corporation 2007. Copyright protects this material. Except as permitted by the Copyright Act 1968 (Cth), reproduction by any means (photocopying, electronic, mechanical, recording or otherwise), making available online, electronic transmission or other publication of this material is prohibited without the prior written permission of the Department of Primary Industries and Fisheries, Queensland and the Fisheries Research and Development Corporation. Inquiries should be addressed to: Intellectual Property and