Modern Pollen from Small Hollows Reflects Athrotaxis Cupressoides
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Vegetation Benchmarks Rainforest and Related Scrub
Vegetation Benchmarks Rainforest and related scrub Eucryphia lucida Vegetation Condition Benchmarks version 1 Rainforest and Related Scrub RPW Athrotaxis cupressoides open woodland: Sphagnum peatland facies Community Description: Athrotaxis cupressoides (5–8 m) forms small woodland patches or appears as copses and scattered small trees. On the Central Plateau (and other dolerite areas such as Mount Field), broad poorly– drained valleys and small glacial depressions may contain scattered A. cupressoides trees and copses over Sphagnum cristatum bogs. In the treeless gaps, Sphagnum cristatum is usually overgrown by a combination of any of Richea scoparia, R. gunnii, Baloskion australe, Epacris gunnii and Gleichenia alpina. This is one of three benchmarks available for assessing the condition of RPW. This is the appropriate benchmark to use in assessing the condition of the Sphagnum facies of the listed Athrotaxis cupressoides open woodland community (Schedule 3A, Nature Conservation Act 2002). Benchmarks: Length Component Cover % Height (m) DBH (cm) #/ha (m)/0.1 ha Canopy 10% - - - Large Trees - 6 20 5 Organic Litter 10% - Logs ≥ 10 - 2 Large Logs ≥ 10 Recruitment Continuous Understorey Life Forms LF code # Spp Cover % Immature tree IT 1 1 Medium shrub/small shrub S 3 30 Medium sedge/rush/sagg/lily MSR 2 10 Ground fern GF 1 1 Mosses and Lichens ML 1 70 Total 5 8 Last reviewed – 2 November 2016 Tasmanian Vegetation Monitoring and Mapping Program Department of Primary Industries, Parks, Water and Environment http://www.dpipwe.tas.gov.au/tasveg RPW Athrotaxis cupressoides open woodland: Sphagnum facies Species lists: Canopy Tree Species Common Name Notes Athrotaxis cupressoides pencil pine Present as a sparse canopy Typical Understorey Species * Common Name LF Code Epacris gunnii coral heath S Richea scoparia scoparia S Richea gunnii bog candleheath S Astelia alpina pineapple grass MSR Baloskion australe southern cordrush MSR Gleichenia alpina dwarf coralfern GF Sphagnum cristatum sphagnum ML *This list is provided as a guide only. -

Edition 2 from Forest to Fjaeldmark the Vegetation Communities Highland Treeless Vegetation
Edition 2 From Forest to Fjaeldmark The Vegetation Communities Highland treeless vegetation Richea scoparia Edition 2 From Forest to Fjaeldmark 1 Highland treeless vegetation Community (Code) Page Alpine coniferous heathland (HCH) 4 Cushion moorland (HCM) 6 Eastern alpine heathland (HHE) 8 Eastern alpine sedgeland (HSE) 10 Eastern alpine vegetation (undifferentiated) (HUE) 12 Western alpine heathland (HHW) 13 Western alpine sedgeland/herbland (HSW) 15 General description Rainforest and related scrub, Dry eucalypt forest and woodland, Scrub, heathland and coastal complexes. Highland treeless vegetation communities occur Likewise, some non-forest communities with wide within the alpine zone where the growth of trees is environmental amplitudes, such as wetlands, may be impeded by climatic factors. The altitude above found in alpine areas. which trees cannot survive varies between approximately 700 m in the south-west to over The boundaries between alpine vegetation communities are usually well defined, but 1 400 m in the north-east highlands; its exact location depends on a number of factors. In many communities may occur in a tight mosaic. In these parts of Tasmania the boundary is not well defined. situations, mapping community boundaries at Sometimes tree lines are inverted due to exposure 1:25 000 may not be feasible. This is particularly the or frost hollows. problem in the eastern highlands; the class Eastern alpine vegetation (undifferentiated) (HUE) is used in There are seven specific highland heathland, those areas where remote sensing does not provide sedgeland and moorland mapping communities, sufficient resolution. including one undifferentiated class. Other highland treeless vegetation such as grasslands, herbfields, A minor revision in 2017 added information on the grassy sedgelands and wetlands are described in occurrence of peatland pool complexes, and other sections. -

Kosipe Revisited
Peat in the mountains of New Guinea G.S. Hope Department of Archaeology and Natural History, Australian National University, Canberra, Australia _______________________________________________________________________________________ SUMMARY Peatlands are common in montane areas above 1,000 m in New Guinea and become extensive above 3,000 m in the subalpine zone. In the montane mires, swamp forests and grass or sedge fens predominate on swampy valley bottoms. These mires may be 4–8 m in depth and up to 30,000 years in age. In Papua New Guinea (PNG) there is about 2,250 km2 of montane peatland, and Papua Province (the Indonesian western half of the island) probably contains much more. Above 3,000 m, peat soils form under blanket bog on slopes as well as on valley floors. Vegetation types include cushion bog, grass bog and sedge fen. Typical peat depths are 0.5‒1 m on slopes, but valley floors and hollows contain up to 10 m of peat. The estimated total extent of mountain peatland is 14,800 km2 with 5,965 km2 in PNG and about 8,800 km2 in Papua Province. The stratigraphy, age structure and vegetation histories of 45 peatland or organic limnic sites above 750 m have been investigated since 1965. These record major vegetation shifts at 28,000, 17,000‒14,000 and 9,000 years ago and a variable history of human disturbance from 14,000 years ago with extensive clearance by the mid- Holocene at some sites. While montane peatlands were important agricultural centres in the Holocene, the introduction of new dryland crops has resulted in the abandonment of some peatlands in the last few centuries. -

Alpine Sphagnum Bogs and Associated Fens
Alpine Sphagnum Bogs and Associated Fens A nationally threatened ecological community Environment Protection and Biodiversity Conservation Act 1999 Policy Statement 3.16 This brochure is designed to assist land managers, owners and occupiers to identify, assess and manage the Alpine Sphagnum Bogs and Associated Fens, an ecological community listed under Australia’s national environment law, the Environment Protection and Biodiversity Conservation Act 1999 (EPBC Act). The brochure is a companion document to the listing advice which can be found at the Australian Government’s Species Profile and Threats Database (SPRAT). Please go to the Alpine Sphagnum Bogs and Associated Fens ecological community profile in SPRAT, then click on the ‘Details’ link: www.environment.gov.au/cgi-bin/sprat/public/publiclookupcommunities.pl • The Alpine Sphagnum Bogs and Associated Fens ecological community is found in small pockets in the high country of Tasmania, Victoria, New South Wales and the Australian Capital Territory. • The Alpine Sphagnum Bogs and Associated Fens ecological community can usually be defined by the presence or absence of sphagnum moss. • Long term conservation and restoration of this ecological community is essential in order to protect vital inland water resources. • Implementing favourable land use and management practices is encouraged at sites containing this ecological community. Disclaimer The contents of this document have been compiled using a range of source materials. This document is valid as at August 2009. The Commonwealth Government is not liable for any loss or damage that may be occasioned directly or indirectly through the use of or reliance on the contents of the document. © Commonwealth of Australia 2009 This work is copyright. -

Plant Life of Western Australia
INTRODUCTION The characteristic features of the vegetation of Australia I. General Physiography At present the animals and plants of Australia are isolated from the rest of the world, except by way of the Torres Straits to New Guinea and southeast Asia. Even here adverse climatic conditions restrict or make it impossible for migration. Over a long period this isolation has meant that even what was common to the floras of the southern Asiatic Archipelago and Australia has become restricted to small areas. This resulted in an ever increasing divergence. As a consequence, Australia is a true island continent, with its own peculiar flora and fauna. As in southern Africa, Australia is largely an extensive plateau, although at a lower elevation. As in Africa too, the plateau increases gradually in height towards the east, culminating in a high ridge from which the land then drops steeply to a narrow coastal plain crossed by short rivers. On the west coast the plateau is only 00-00 m in height but there is usually an abrupt descent to the narrow coastal region. The plateau drops towards the center, and the major rivers flow into this depression. Fed from the high eastern margin of the plateau, these rivers run through low rainfall areas to the sea. While the tropical northern region is characterized by a wet summer and dry win- ter, the actual amount of rain is determined by additional factors. On the mountainous east coast the rainfall is high, while it diminishes with surprising rapidity towards the interior. Thus in New South Wales, the yearly rainfall at the edge of the plateau and the adjacent coast often reaches over 100 cm. -

The Ecology and Biogeography of Athrotaxis D. Don
THE ECOLOGY AND BIOGEOGRAPHY OF ATHROTAXIS D. DON. ,smo,e5 by Philip J. Cullen B.Sc. (Forestry) submitted in fulfilment of the requirements for the degree of Master of Science UNIVERSITY OF TASMANIA HOBART February 1987 DECLARATION This thesis contains no material which has been submitted for the award of any other degree or diploma in any university and contains no copy or paraphrases of material previously published or writtern by another person, except where due reference is made in the text. P. J. Cullen. CONTENTS PAGE ACKNOWLEGEMENTS ABSTRACT II LIST OF FIGURES IV LIST OF TABLES VI LIST OF PLATES VIII CHAPTER 1: INTRODUCTION , 1 CHAPTER 2: DISTRIBUTION AND SYNECOLOGY 2.1 Introduction 8 2.2 Methods 14 2.3 Results and Discussion 2.3.1 Stand Classification 19 2.3.2 Stand Ordination 32 CHAPTER 3: THE REGENERATION MODES OF ATHROTAXIS CUPRESSOIDES AND ATHROTAXIS SELAGINOIDES 3.1 Introduction 38 3.2 Methods 3.2.1 Stand demography 40 3.2.2 Spatial distribution of seedlings 43 3.2.3 Seed dispersal 44 3.2.4 Vegetative reproduction 45 3.3 Results and Discussion 3.3.1 Size/age correlations 48 3.3.2 Seed production and dispersal 52 3.3.3 Vegetative regeneration 55 3.3.4 Stand demography and seedling distribution of Athrotaxis cupressoides 60 3.3.5 Stand demography and seedling distribution of Athrotaxis seiaginoides 82 3.3.6 The presence of Athrotaxis laxifolia . 88 3.4 Conclusion 89 CHAPTER THE RELATIVE FROST RESISTANCE OF ATHROTAXIS CUPRESSOIDES AND ATHROTAXIS SELAGINOIDES SEEDLINGS 4.1 Introduction 92 4.2 Methods 93 4,3 Results 96 4.4 Discussion 99 CHAPTER 5: THE EFFECT OF GRAZING ON THE SEEDLING REGENERATION OF ATHROTAXIS CUPRESSOIDES 5.1 Introduction 107 5.2 Methods 107 5.3 Results 109 5.4 Discussion 111 CHAPTER 6: DISCUSSION 116 REFERENCES 135 APPENDIX A: REGENERATION PATTERNS IN POPULATIONS OF ATHROTAXIS SELAGINOIDES D. -

Alternative Translation Initiation Codons for the Plastid Maturase Matk: Unraveling the Pseudogene Misconception in the Orchidaceae Michelle M
Barthet et al. BMC Evolutionary Biology (2015) 15:210 DOI 10.1186/s12862-015-0491-1 RESEARCH ARTICLE Open Access Alternative translation initiation codons for the plastid maturase MatK: unraveling the pseudogene misconception in the Orchidaceae Michelle M. Barthet1,2* , Keenan Moukarzel3, Kayla N. Smith1, Jaimin Patel3 and Khidir W. Hilu3 Abstract Background: The plastid maturase MatK has been implicated as a possible model for the evolutionary “missing link” between prokaryotic and eukaryotic splicing machinery. This evolutionary implication has sparked investigations concerning the function of this unusual maturase. Intron targets of MatK activity suggest that this is an essential enzyme for plastid function. The matK gene, however, is described as a pseudogene in many photosynthetic orchid species due to presence of premature stop codons in translations, and its high rate of nucleotide and amino acid substitution. Results: Sequence analysis of the matK gene from orchids identified an out-of-frame alternative AUG initiation codon upstream from the consensus initiation codon used for translation in other angiosperms. We demonstrate translation from the alternative initiation codon generates a conserved MatK reading frame. We confirm that MatK protein is expressed and functions in sample orchids currently described as having a matK pseudogene using immunodetection and reverse-transcription methods. We demonstrate using phylogenetic analysis that this alternative initiation codon emerged de novo within the Orchidaceae, with several reversal events at the basal lineage and deep in orchid history. Conclusion: These findings suggest a novel evolutionary shift for expression of matK in the Orchidaceae and support the function of MatK as a group II intron maturase in the plastid genome of land plants including the orchids. -

Tasmania - from the Wet West to the Dry East
This collection is maintained with the assistance of the Tasmania - from the wet west to the dry east. Regional Branch of the Australian Plant Society. Influences on the development of the Tasmanian plant mix Montane moorland and cool oceanic heathland When Gondwana existed as a super Geology of Tasmania Vegetation Map of Tasmania The Tasmanian highland vegetation developed in isolation from the Australian Alps. Even during ice continent, Australia and Tasmania, Africa, ages, hundreds of kilometres of lowland vegetation separated the two high altitude environments. South America, New Zealand and Antarctica shared many plant families and some Montane plants have to cope with wide temperature fluctuations, with periods of below 0°C and Genera. exposure to winds. Cold may be prolonged if the ground freezes. Plants may be blanketed by snow or BASS STRAIT the mountains by cloud. Snowmelt or clear weather can cause intense rays of light, resulting in high For example, the protea family has members temperature. Wind or sun can dry the plant and soil. in all those land masses except Antarctica. The Southern Africa panel covers the protea family more fully. Plants require moisture and warmth. Small hard leaves offer Tasmania was the last land mass to break protection from the drying away from Antarctica. The opening of the effects of sun and wind. Low gap between these land masses allowed the ocean to circulate growth avoids wind. Branches around Antarctica, cooling the earth’s climate and so locking up grow close together to shelter vast quantities of water as ice. the parts of each plant. -

World Heritage Values and to Identify New Values
FLORISTIC VALUES OF THE TASMANIAN WILDERNESS WORLD HERITAGE AREA J. Balmer, J. Whinam, J. Kelman, J.B. Kirkpatrick & E. Lazarus Nature Conservation Branch Report October 2004 This report was prepared under the direction of the Department of Primary Industries, Water and Environment (World Heritage Area Vegetation Program). Commonwealth Government funds were contributed to the project through the World Heritage Area program. The views and opinions expressed in this report are those of the authors and do not necessarily reflect those of the Department of Primary Industries, Water and Environment or those of the Department of the Environment and Heritage. ISSN 1441–0680 Copyright 2003 Crown in right of State of Tasmania Apart from fair dealing for the purposes of private study, research, criticism or review, as permitted under the Copyright Act, no part may be reproduced by any means without permission from the Department of Primary Industries, Water and Environment. Published by Nature Conservation Branch Department of Primary Industries, Water and Environment GPO Box 44 Hobart Tasmania, 7001 Front Cover Photograph: Alpine bolster heath (1050 metres) at Mt Anne. Stunted Nothofagus cunninghamii is shrouded in mist with Richea pandanifolia scattered throughout and Astelia alpina in the foreground. Photograph taken by Grant Dixon Back Cover Photograph: Nothofagus gunnii leaf with fossil imprint in deposits dating from 35-40 million years ago: Photograph taken by Greg Jordan Cite as: Balmer J., Whinam J., Kelman J., Kirkpatrick J.B. & Lazarus E. (2004) A review of the floristic values of the Tasmanian Wilderness World Heritage Area. Nature Conservation Report 2004/3. Department of Primary Industries Water and Environment, Tasmania, Australia T ABLE OF C ONTENTS ACKNOWLEDGMENTS .................................................................................................................................................................................1 1. -

Supporting Information
Supporting Information Mao et al. 10.1073/pnas.1114319109 SI Text BEAST Analyses. In addition to a BEAST analysis that used uniform Selection of Fossil Taxa and Their Phylogenetic Positions. The in- prior distributions for all calibrations (run 1; 144-taxon dataset, tegration of fossil calibrations is the most critical step in molecular calibrations as in Table S4), we performed eight additional dating (1, 2). We only used the fossil taxa with ovulate cones that analyses to explore factors affecting estimates of divergence could be assigned unambiguously to the extant groups (Table S4). time (Fig. S3). The exact phylogenetic position of fossils used to calibrate the First, to test the effect of calibration point P, which is close to molecular clocks was determined using the total-evidence analy- the root node and is the only functional hard maximum constraint ses (following refs. 3−5). Cordaixylon iowensis was not included in in BEAST runs using uniform priors, we carried out three runs the analyses because its assignment to the crown Acrogymno- with calibrations A through O (Table S4), and calibration P set to spermae already is supported by previous cladistic analyses (also [306.2, 351.7] (run 2), [306.2, 336.5] (run 3), and [306.2, 321.4] using the total-evidence approach) (6). Two data matrices were (run 4). The age estimates obtained in runs 2, 3, and 4 largely compiled. Matrix A comprised Ginkgo biloba, 12 living repre- overlapped with those from run 1 (Fig. S3). Second, we carried out two runs with different subsets of sentatives from each conifer family, and three fossils taxa related fi to Pinaceae and Araucariaceae (16 taxa in total; Fig. -

Biogeography of the Monocotyledon Astelioid Clade (Asparagales): a History of Long-Distance Dispersal and Diversification with Emerging Habitats
Zurich Open Repository and Archive University of Zurich Main Library Strickhofstrasse 39 CH-8057 Zurich www.zora.uzh.ch Year: 2021 Biogeography of the monocotyledon astelioid clade (Asparagales): A history of long-distance dispersal and diversification with emerging habitats Birch, Joanne L ; Kocyan, Alexander Abstract: The astelioid families (Asteliaceae, Blandfordiaceae, Boryaceae, Hypoxidaceae, and Lanari- aceae) have centers of diversity in Australasia and temperate Africa, with secondary centers of diversity in Afromontane Africa, Asia, and Pacific Islands. The global distribution of these families makes this an excellent lineage to test if current distribution patterns are the result of vicariance or long-distance dispersal and to evaluate the roles of tertiary climatic and geological drivers in lineage diversification. Sequence data were generated from five chloroplast regions (petL-psbE, rbcL, rps16-trnK, trnL-trnLF, trnS-trnSG) for 104 ingroup species sampled across global diversity. The astelioid phylogeny was inferred using maximum parsimony, maximum likelihood, and Bayesian inference methods. Divergence dates were estimated with a relaxed clock applied in BEAST. Ancestral ranges were reconstructed in ’BioGeoBEARS’ applying the corrected Akaike information criterion to test for the best-fit biogeographic model. Diver- sification rates were estimated in Bayesian Analysis of Macroevolutionary Mixtures [BAMM]. Astelioid relationships were inferred as Boryaceae(Blandfordiaceae(Asteliaceae(Hypoxidaceae plus Lanariaceae))). The crown astelioid node was dated to the Late Cretaceous (75.2 million years; 95% highest posterior densities interval 61.0-90.0 million years) with an inferred Eastern Gondwanan origin. However, aste- lioid speciation events have not been shaped by Gondwanan vicariance. Rather long-distance dispersal since the Eocene is inferred to account for current distributions. -
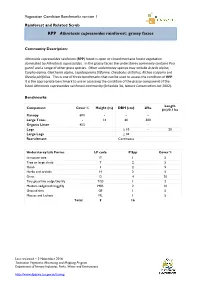
RPP Athrotaxis Cupressoides Rainforest: Grassy Facies
Vegetation Condition Benchmarks version 1 Rainforest and Related Scrub RPP Athrotaxis cupressoides rainforest: grassy facies Community Description: Athrotaxis cupressoides rainforest (RPP) forest is open or closed montane forest vegetation dominated by Athrotaxis cupressoides. In the grassy facies the understorey commonly contains Poa gunnii and a range of other grass species. Other understorey species may include Astelia alpina, Carpha alpina, Gleichenia alpina, Lepidosperma filiforme, Oreobolus distichus, Richea scoparia and Olearia pinifolius. This is one of three benchmarks that can be used to assess the condition of RPP. It is the appropriate benchmark to use in assessing the condition of the grassy component of the listed Athrotaxis cupressoides rainforest community (Schedule 3A, Nature Conservation Act 2002). Benchmarks: Length Component Cover % Height (m) DBH (cm) #/ha (m)/0.1 ha Canopy 60% - - - Large Trees - 14 60 300 Organic Litter 45% - Logs ≥ 10 - 20 Large Logs ≥ 30 Recruitment Continuous Understorey Life Forms LF code # Spp Cover % Immature tree IT 1 5 Tree or large shrub T 2 5 Shrub S 2 5 Herbs and orchids H 2 5 Grass G 4 20 Tiny grass/tiny sedge/tiny lily TGS 1 2 Medium sedge/rush/sagg/lily MSR 2 10 Ground fern GF 1 5 Mosses and Lichens ML 1 5 Total 9 16 Last reviewed – 2 November 2016 Tasmanian Vegetation Monitoring and Mapping Program Department of Primary Industries, Parks, Water and Environment http://www.dpipwe.tas.gov.au/tasveg RPP Athrotaxis cupressoides rainforest: grassy facies Species lists: Canopy Tree Species Common Name Notes Athrotaxis cupressoides pencil pine Cradle Mountain area; some Athrotaxis selaginoides king billy pine closed canopy forests Cradle Mountain area; some Nothofagus cunninghamii myrtle beech closed canopy forests Typical Understorey Species * Common Name LF Code Nothofagus cunninghamii myrtle beech T Phyllocladus aspleniifolius celery top pine T Olearia pinifolia prickly daisybush S Richea species richea S Poa gunnii gunns snowgrass G Rytidosperma spp.