Localization of Glucocorticoid Receptor Interacting Protein 1 in Murine
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Urology & Incontinence
Urology & Incontinence - Glossary of Terms Anti-reflux Refers to a tube or collapsible material within a urine collection device to help prevent urine from reentering the tubing. Applicator collar Found on some Hollister male external catheters, a plastic guide with notches for the thumb and forefinger to assist proper placement against the tip of the penis. Aseptic intermittent catheterization The process of performing intermittent catheterization using sterile equipment and inserting the catheter in a sterile way. This would include a sterile ready-to-use product that can be inserted with gloves using a no-touch technique (e.g., the Advance Plus intermittent catheter or a VaPro hydrophilic catheter). Benzalkonium chloride (BZK) An antimicrobial solution used for cleansing the urethral opening prior to inserting an intermittent catheter. Does not stain skin or clothing. Bladder A collapsible balloon-like muscular organ that lies in the pelvis and functions to store and expel urine. Bladder catheterization A procedure in which a catheter is passed through the urethra or stoma into the bladder, usually for the purpose of draining urine. Bladder control The ability to control urination. Bladder diary A printed or electronic form to keep track of when one urinates or leaks urine. Catheter (urinary) A special type of hollow tube inserted through the urethra or a stoma to the bladder to withdraw urine or instill medication. Catheterization The process of inserting a tube into the bladder to drain urine. Clean intermittent catheterization The process of emptying the bladder using a clean intermittent catheter. It involves inserting and removing a catheter, typically several times a day. -
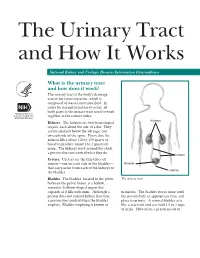
The Urinary Tract and How It Works
The Urinary Tract and How It Works National Kidney and Urologic Diseases Information Clearinghouse What is the urinary tract and how does it work? The urinary tract is the body’s drainage system for removing urine, which is composed of wastes and extra fluid. In order for normal urination to occur, all body parts in the urinary tract need to work together in the correct order. Kidneys Kidneys. The kidneys are two bean-shaped organs, each about the size of a fist. They are located just below the rib cage, one on each side of the spine. Every day, the kidneys filter about 120 to 150 quarts of blood to produce about 1 to 2 quarts of urine. The kidneys work around the clock; a person does not control what they do. Ureters Ureters. Ureters are the thin tubes of muscle—one on each side of the bladder— Bladder that carry urine from each of the kidneys to Urethra the bladder. Bladder. The bladder, located in the pelvis The urinary tract between the pelvic bones, is a hollow, muscular, balloon-shaped organ that expands as it fills with urine. Although a urination. The bladder stores urine until person does not control kidney function, the person finds an appropriate time and a person does control when the bladder place to urinate. A normal bladder acts empties. Bladder emptying is known as like a reservoir and can hold 1.5 to 2 cups of urine. How often a person needs to urinate depends on how quickly the kidneys Why is the urinary tract produce the urine that fills the bladder. -

15-1040-Junu Oh-Neuronal.Key
Neuronal Control of the Bladder Seung-June Oh, MD Department of urology, Seoul National University Hospital Seoul National University College of Medicine Contents Relevant end organs and nervous system Reflex pathways Implication in the sacral neuromodulation Urinary bladder ! body: detrusor ! trigone and bladder neck Urethral sphincters B Preprostatic S Smooth M. Sphincter Passive Prostatic S Skeletal M. Sphincter P Prostatic SS P-M Striated Sphincter Membraneous SS Periurethral Striated M. Pubococcygeous Spinal cord ! S2–S4 spinal cord ! primary parasympathetic micturition center ! bladder and distal urethral sphincter ! T11-L2 spinal cord ! sympathetic outflow ! bladder and proximal urethral sphincter Peripheral innervation ! The lower urinary tract is innervated by 3 principal sets of peripheral nerves: ! parasympathetic -pelvic n. ! sympathetic-hypogastric n. ! somatic nervous systems –pudendal n. ! Parasympathetic and sympathetic nervous systems form pelvic plexus at the lateral side of the rectum before reaching bladder and sphincter Sympathetic & parasympathetic systems ! Sympathetic pathways ! originate from the T11-L2 (sympathetic nucleus; intermediolateral column of gray matter) ! inhibiting the bladder body and excite the bladder base and proximal urethral sphincter ! Parasympathetic nerves ! emerge from the S2-4 (parasympathetic nucleus; intermediolateral column of gray matter) ! exciting the bladder and relax the urethra Sacral somatic system !emerge from the S2-4 (Onuf’s nucleus; ventral horn) !form pudendal nerve, providing -

Nerve Disease and Bladder Control
Nerve Disease and Bladder Control National Kidney and Urologic Diseases Information Clearinghouse For the urinary system to do its job, muscles and nerves must work together to hold Brain urine in the bladder and then release it at the right time. Nerves carry messages from NATIONAL the bladder to the brain to let it know when INSTITUTES the bladder is full. They also carry messages OF HEALTH Central nervous from the brain to the bladder, telling muscles system (brain either to tighten or release. A nerve prob and spinal cord) lem might affect your bladder control if the nerves that are supposed to carry messages Spinal cord between the brain and the bladder do not work properly. Nerve signals U.S. Department to bladder of Health and Bladder and sphincter Human Services What bladder control muscles problems does nerve damage cause? Nerves that work poorly can lead to three Urethra different kinds of bladder control problems. Overactive bladder. Damaged nerves may Sphincter muscles send signals to the bladder at the wrong time, causing its muscles to squeeze with Nerves carry signals from the brain to the bladder out warning. The symptoms of overactive and sphincter. bladder include • urinary frequency—defined as urination eight or more times a day or two or nerves to the sphincter muscles are dam more times at night aged, the muscles may become loose and allow leakage or stay tight when you are • urinary urgency—the sudden, strong trying to release urine. need to urinate immediately Urine retention. For some people, nerve • urge incontinence—leakage of urine damage means their bladder muscles do that follows a sudden, strong urge to not get the message that it is time to release urinate urine or are too weak to completely empty Poor control of sphincter muscles. -
Laboratory 8 - Urinary and Reproductive Systems
Laboratory 8 - Urinary and Reproductive Systems Urinary System Please read before starting: It is easy to damage the structures of the reproductive system as you expose structures associated with excretion, so exercise caution as you do this. Please also note that we will have drawings available as well to help you find and identify the structures described below. The major blood vessels serving the kidneys are the Renal renal artery and the renal pyramid vein., which are located deep in the parietal peritoneum. The renal artery is a branch of the dorsal aorta that comes off Renal further caudal than the cranial pelvis mesenteric artery. Dissect the left kidney in situ, dividing it into dorsal and ventral portions by making a frontal section along the outer periphery. Observe the renal cortex renal medulla (next layer in) renal pyramids renal pelvis ureter (see above diagram) The kidneys include a variety of structures including an arterial supply, a venous return, extensive capillary networks around each nephron and then, of course, the filtration and reabsorption apparatus. These structures are primarily composed of nephrons (the basic functional unit of the kidney) and the ducts which carry urine away from the nephron (the collecting ducts and larger ducts eventually draining these into the ureters from each kidney. The renal pyramids contain the extensions of the nephrons into the renal medulla (the Loops of Henle) and the collecting ducts. Urine is eventually emptied into the renal pelvis before leaving the kidneys in the ureters. The ureters leaves the kidneys medially at approximately the midpoint of the organs and then run caudal to the urinary bladder. -

Incomplete Bladder Emptying and Tips to Help
A guide for patients about incomplete bladder emptying and tips to help What is incomplete bladder emptying? When the bladder doesn't empty properly this may cause numerous problems. One of the problems is known as overflow incontinence. This is when the bladder doesn’t empty and urine leaks out. You may not get the message to go to the toilet either. The bladder never empties completely so some residue is normal. You may find it difficult to start to pass water and that even when you have started; the flow is weak and slow. You might find that you dribble after you have finished passing water. Perhaps you dribble urine all the time, even without noticing. What causes incomplete bladder emptying? Incomplete bladder emptying occurs when the muscles of the bladder are not able to squeeze properly to empty the bladder. This can happen in cases where there may have been nerve or muscle damage, perhaps caused by injury, surgery, or disease such as Parkinson's disease, Multiple Sclerosis and Spina Bifida. Medication can also cause problems with bladder emptying. In some cases, it can be due to an obstruction which is making it more difficult for you to empty your bladder. This obstruction can be caused by an enlarged prostate in men, a kidney stone blocking the urethra, constipation, or stricture of the urethra in either men or women, which makes it difficult for urine to flow out of the bladder outlet. If you are not able to empty completely, your bladder and its muscles may become floppy over time. -

Pre-Clinical Efficacy and Safety Evaluation of Human Amniotic Fluid-Derived Stem Cell Injection in a Mouse Model of Urinary Incontinence
http://dx.doi.org/10.3349/ymj.2015.56.3.648 Original Article pISSN: 0513-5796, eISSN: 1976-2437 Yonsei Med J 56(3):648-657, 2015 Pre-Clinical Efficacy and Safety Evaluation of Human Amniotic Fluid-Derived Stem Cell Injection in a Mouse Model of Urinary Incontinence Jae Young Choi,1* So Young Chun,2* Bum Soo Kim,1 Hyun Tae Kim,1 Eun Sang Yoo,1 Yun-Hee Shon,2 Jeong Ok Lim,2 Seok Joong Yun,3 Phil Hyun Song,4 Sung Kwang Chung,1 James J Yoo,2,5 and Tae Gyun Kwon1,2 1Department of Urology, School of Medicine, Kyungpook National University, Daegu; 2Joint Institute for Regenerative Medicine, Kyungpook National University Hospital, Daegu; 3Department of Urology, College of Medicine, Chungbuk National University, Cheongju; 4Department of Urology, College of Medicine, Yeungnam University, Daegu, Korea; 5Wake Forest Institute for Regenerative Medicine, Wake Forest University School of Medicine, Winston-Salem, NC, USA. Received: March 31, 2014 Purpose: Stem cell-based therapies represent new promises for the treatment of Revised: August 10, 2014 urinary incontinence. This study was performed to assess optimized cell passage Accepted: August 13, 2014 number, cell dose, therapeutic efficacy, feasibility, toxicity, and cell trafficking for Corresponding author: Dr. Tae Gyun Kwon, the first step of the pre-clinical evaluation of human amniotic fluid stem cell (hAF- Department of Urology, School of Medicine, Kyungpook National University, SC) therapy in a urinary incontinence animal model. Materials and Methods: 130 Dongdeok-ro, Jung-gu, The proper cell passage number was analyzed with hAFSCs at passages 4, 6, and Daegu 700-721, Korea. -

Urinary Retention
Urinary Retention National Kidney and Urologic Diseases Information Clearinghouse What is urinary retention? What is the urinary tract Urinary retention is the inability to and how does it work? empty the bladder completely. Urinary The urinary tract is the body’s drainage retention can be acute or chronic. Acute system for removing urine, which is urinary retention happens suddenly and composed of wastes and extra fluid. In lasts only a short time. People with acute order for normal urination to occur, all urinary retention cannot urinate at all, body parts in the urinary tract need to work even though they have a full bladder. together in the correct order. Acute urinary retention, a potentially life-threatening medical condition, Kidneys. The kidneys are two bean-shaped requires immediate emergency treatment. organs, each about the size of a fist. They Acute urinary retention can cause great are located just below the rib cage, one discomfort or pain. on each side of the spine. Every day, the kidneys filter about 120 to 150 quarts of Chronic urinary retention can be a long- blood to produce about 1 to 2 quarts of lasting medical condition. People with urine. The kidneys work around the clock; chronic urinary retention can urinate. a person does not control what they do. However, they do not completely empty all of the urine from their bladders. Ureters. Ureters are the thin tubes of Often people are not even aware they muscle—one on each side of the bladder— have this condition until they develop that carry urine from each of the kidneys to another problem, such as urinary the bladder. -

Male Reproductive Anatomy
Male Reproductive Anatomy Words To Know: Testosterone: Male hormone. Causes hair growth, deeper voice, sperm production. Bladder: Organ that holds urine until excreted from the body. Testicle / Testes: Male organ that produces the sperm. 100 million sperm produced a day. The testes are developed high in the body and nerve endings are still there. Sperm Cells: Male reproductive cell produced in the testes. Scrotum: This is a sac that regulates the temperature of the testes. Keeps temperature at 95℉ and will rise closer to the body (to get warmer, use body heat) and lower (cool down) to help with this. Epididymis: Sperm is stored here. Can hold sperm for 6 weeks or until mature. Vas Deferens: A tube that travels from the epididymis to the urethra. Sperm travel along this tube. Seminal Vesicle: This vesicle adds a sugary fluid to the semen. Prostate Gland: This gland adds a chemical fluid to the semen. Cowper’s Gland: Secretes a fluid that helps make the environment safer for the sperm. Urethra: Brings urine and semen out of the body. Penis: Is made up of a spongy tissue. Erection: When the penis becomes engorged with blood and hard. Semen: Fluid ejaculated from the penis. Six Ways to Care for the Male Reproductive System! 1. Choose Abstinence. Abstinence is choosing not to be sexually active. Choosing abstinence prevents infection with sexually transmitted diseases. It also keeps you from becoming a teen father. 2. Have Regular Medical Checkups. Your physician can examine you and discuss body changes. 3. Bathe or Shower Daily. Keep your external reproductive organs clean. -

Ureter Urinary Bladder Seminal Vesicle Ampulla of Ductus Deferens
Ureter Urinary bladder Seminal vesicle Prostatic urethra Ampulla of Pubis ductus deferens Membranous urethra Ejaculatory duct Urogenital diaphragm Rectum Erectile tissue Prostate of the penis Bulbo-urethral gland Spongy urethra Shaft of the penis Ductus (vas) deferens Epididymis Glans penis Testis Prepuce Scrotum External urethral (a) orifice © 2018 Pearson Education, Inc. 1 Urinary bladder Ureter Ampulla of ductus deferens Seminal vesicle Ejaculatory Prostate duct Prostatic Bulbourethral urethra gland Membranous Ductus urethra deferens Root of penis Erectile tissues Epididymis Shaft (body) of penis Testis Spongy urethra Glans penis Prepuce External urethral (b) orifice © 2018 Pearson Education, Inc. 2 Spermatic cord Blood vessels and nerves Seminiferous tubule Rete testis Ductus (vas) deferens Lobule Septum Tunica Epididymis albuginea © 2018 Pearson Education, Inc. 3 Seminiferous tubule Basement membrane Spermatogonium 2n 2n Daughter cell (stem cell) type A (remains at basement Mitosis 2n membrane as a stem cell) Growth Daughter cell type B Enters (moves toward tubule prophase of lumen) meiosis I 2n Primary spermatocyte Meiosis I completed Meiosis n n Secondary spermatocytes Meiosis II n n n n Early spermatids n n n n Late spermatids Spermatogenesis Spermiogenesis Sperm n n n n Lumen of seminiferous tubule © 2018 Pearson Education, Inc. 4 Gametes (n = 23) n Egg n Sperm Meiosis Fertilization Multicellular adults Zygote 2n (2n = 46) (2n = 46) Mitosis and development © 2018 Pearson Education, Inc. 5 Provides genetic Provides instructions and a energy for means of penetrating mobility the follicle cell capsule and Plasma membrane oocyte membrane Neck Provides Tail for mobility Head Midpiece Axial filament Acrosome of tail Nucleus Mitochondria Proximal centriole (b) © 2018 Pearson Education, Inc. -

The Length of the Male Urethra International Braz J Urol Vol
Clinical Urology The Length of the Male Urethra International Braz J Urol Vol. 34 (4): 451-456, July - August, 2008 The Length of the Male Urethra Tobias. S. Kohler, Mitchell Yadven, Ankur Manvar, Nathan Liu, Manoj Monga Division of Urology (TSK), Southern Illinois University, Springfield, IL, Department of Urologic Surgery (MY, NL, MM), University of Minnesota, Minneapolis, MN, Department of Urologic Surgery (AM), Northwestern University, Feinberg School of Medicine, Chicago, Illinois, USA ABSTRACT Purpose: Catheter-based medical devices are an important component of the urologic armamentarium. To our knowledge, there is no population-based data regarding normal male urethral length. We evaluated the length of the urethra in men with normal genitourinary anatomy undergoing either Foley catheter removal or standard cystoscopy. Materials and Methods: Male urethral length was obtained in 109 men. After study permission was obtained, the subject’s penis was placed on a gentle stretch and the catheter was marked at the tip of the penis. The catheter was then removed and the distance from the mark to the beginning of the re-inflated balloon was measured. Alternatively, urethral length was measured at the time of cystoscopy, on removal of the cystoscope. Data on age, weight, and height was obtained in patients when possible. Results: The mean urethral length was 22.3 cm with a standard deviation of 2.4 cm. Urethral length varied between 15 cm and 29 cm. No statistically significant correlation was found between urethral length and height, weight, body mass index (BMI), or age. Conclusions: Literature documenting the length of the normal male adult urethra is scarce. -
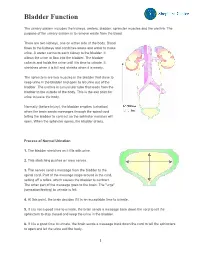
Bladder Function
Bladder Function The urinary system includes the kidneys, ureters, bladder, sphincter muscles and the urethra. The purpose of the urinary system is to remove waste from the blood. There are two kidneys, one on either side of the body. Blood flows to the kidneys and combines waste and water to make urine. A ureter connects each kidney to the bladder. It allows the urine to flow into the bladder. The bladder collects and holds the urine until it is time to urinate. It stretches when it is full and shrinks when it is empty. The sphincters are two muscles in the bladder that close to keep urine in the bladder and open to let urine out of the bladder. The urethra is a muscular tube that leads from the bladder to the outside of the body. This is the exit point for urine to leave the body. Normally (before injury), the bladder empties (urination) when the brain sends messages through the spinal cord telling the bladder to contract so the sphincter muscles will open. When the sphincter opens, the bladder drains. Process of Normal Urination 1. The bladder stretches as it fills with urine. 2. This stretching pushes on area nerves. 3. The nerves send a message from the bladder to the spinal cord. Part of the message loops around in the cord, setting off a reflex, which causes the bladder to contract. The other part of the message goes to the brain. The "urge" (sensation/feeling) to urinate is felt. 4. At this point, the brain decides if it is an acceptable time to urinate.