Fossil Magnoliaceae: a Review of Literature
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Recommended Trees for Winnetka
RECOMMENDED TREES FOR WINNETKA SHADE TREES Common_Name Scientific_Name Ohio Buckeye Acer galbra Miyabe Maple Acer miyabei Black Maple Acer nigrum Norway Maple Acer plantanoides v. ___ Sugar Maple (many cultivars) Acer saccharum Shangtung Maple Acer truncatum Autumn Blaze or Marmo Maple Acer x freemanii Red Horsechestnut Aesculus x carnea 'Briotii' Horsechestnut Aesulus hippocastanum Alder Alnus glutinosa Yellowwood Caldrastis lutea Upright European Hornbeam Carpinus betulus “Fastigata” American Hornbeam Carpinus carolinians Hickory Carya ovata Catalpa Catalpa speciosa Hackberry Celtis occidentalis Katsuratree Cercidiphyllum japonicum Turkish Filbert Corylus colurna American Beech Fagus grandifolia Green Beech Fagus sylvatica European Beech Fagus sylvatica Ginkgo Ginkgo biloba Thornless Honeylocust Gleditsia triacanthos inermis Kentucky Coffeetree Gymnocladus dioica Goldenraintree Koelreuteria paniculata Sweetgum Liquidambar styraciflua Tulip Tree Liriodendron tulipfera Black gum, Tupelo Liriodendron tulipfera Hophornbeam Ostrya virginiana Corktree Phellodendron amurense Exclamation Plantree Plantanus x aceerifolia Quaking Aspen Populus tremuloides Swamp White Oak Quercus bicolor Shingle Oak Quercus imbricaria Bur Oak Quercus macrocarpa Chinkapin Oak Quercus muehlenbergii English Oak Quercus robur Red Oak Quercus rubra Schumard Oak Quercus shumardii Black Oak Quercus velutina May 2015 SHADE TREES Common_Name Scientific_Name Sassafras Sassafras albidum American Linden Tilia Americana Littleleaf Linden (many cultivars) Tilia cordata Silver -

Beyond the Fringe
Beyond the Fringe By Margaret Klein Wilson, Fairfax Master Gardener Intern The final flourish of native spring flowering trees belongs to fringe trees (Chionanthus virginicus). The measured parade of seasonal color — dogwoods and redbuds, flowering cherry and plum — is neatly capped by the fluttering grace of abundant white blossoms that engulf this small, often edge habitat tree beginning in mid-May. Encountering a fringe tree on a breezy spring day is to see masses of white corollas of petals casting their light perfume across the landscape. Is it any wonder Linnaeus named Yale ©2021 Copyright and noted this genus Chionanthus (chion = University snow; anthos = flower) as he compiled the photo: Genera Plantarum (1753)? Less poetically, fringe tree’s common names include Old Man’s Beard, Grancy graybeard, flowering ash and white fringe tree. Genus Chionanthus is a member of the Olive Family, the Olaceae, which includes 29 genera and over 600 species of trees and shrubs common in southeastern Asia and Australasia. Trees in this genus have uses both practical and ornamental: olive for food, ash for its lumber, and forsythia, gardenia and privets for sheer domestic beauty. In North America, ash trees in the genus Fraxinus, devilwood (Osmantus americanus) and Forestiera (swampprivet) are fringe tree’s near relatives. Only two species of fringe tree exist: C. virginicus in eastern North America and C. retusus, native to China. In his citation naming fringe tree the Virginia Native Plant Society 1997 Wildflower of the Year, C. F. Saachi explains this geographic disconnect: “This unusual biogeographic pattern, with different species within a genus … separated by several thousand miles, is a product of major geologic events including mountain building and the effects of glaciation. -

Indiana's Native Magnolias
FNR-238 Purdue University Forestry and Natural Resources Know your Trees Series Indiana’s Native Magnolias Sally S. Weeks, Dendrologist Department of Forestry and Natural Resources Purdue University, West Lafayette, IN 47907 This publication is available in color at http://www.ces.purdue.edu/extmedia/fnr.htm Introduction When most Midwesterners think of a magnolia, images of the grand, evergreen southern magnolia (Magnolia grandiflora) (Figure 1) usually come to mind. Even those familiar with magnolias tend to think of them as occurring only in the South, where a more moderate climate prevails. Seven species do indeed thrive, especially in the southern Appalachian Mountains. But how many Hoosiers know that there are two native species Figure 2. Cucumber magnolia when planted will grow well throughout Indiana. In Charles Deam’s Trees of Indiana, the author reports “it doubtless occurred in all or nearly all of the counties in southern Indiana south of a line drawn from Franklin to Knox counties.” It was mainly found as a scattered, woodland tree and considered very local. Today, it is known to occur in only three small native populations and is listed as State Endangered Figure 1. Southern magnolia by the Division of Nature Preserves within Indiana’s Department of Natural Resources. found in Indiana? Very few, I suspect. No native As the common name suggests, the immature magnolias occur further west than eastern Texas, fruits are green and resemble a cucumber so we “easterners” are uniquely blessed with the (Figure 3). Pioneers added the seeds to whisky presence of these beautiful flowering trees. to make bitters, a supposed remedy for many Indiana’s most “abundant” species, cucumber ailments. -

THE Magnoliaceae Liriodendron L. Magnolia L
THE Magnoliaceae Liriodendron L. Magnolia L. VEGETATIVE KEY TO SPECIES IN CULTIVATION Jan De Langhe (1 October 2014 - 28 May 2015) Vegetative identification key. Introduction: This key is based on vegetative characteristics, and therefore also of use when flowers and fruits are absent. - Use a 10× hand lens to evaluate stipular scars, buds and pubescence in general. - Look at the entire plant. Young specimens, shade, and strong shoots give an atypical view. - Beware of hybridisation, especially with plants raised from seed other than wild origin. Taxa treated in this key: see page 10. Questionable/frequently misapplied names: see page 10. Names referred to synonymy: see page 11. References: - JDL herbarium - living specimens, in various arboreta, botanic gardens and collections - literature: De Meyere, D. - (2001) - Enkele notities omtrent Liriodendron tulipifera, L. chinense en hun hybriden in BDB, p.23-40. Hunt, D. - (1998) - Magnolias and their allies, 304p. Bean, W.J. - (1981) - Magnolia in Trees and Shrubs hardy in the British Isles VOL.2, p.641-675. - or online edition Clarke, D.L. - (1988) - Magnolia in Trees and Shrubs hardy in the British Isles supplement, p.318-332. Grimshaw, J. & Bayton, R. - (2009) - Magnolia in New Trees, p.473-506. RHS - (2014) - Magnolia in The Hillier Manual of Trees & Shrubs, p.206-215. Liu, Y.-H., Zeng, Q.-W., Zhou, R.-Z. & Xing, F.-W. - (2004) - Magnolias of China, 391p. Krüssmann, G. - (1977) - Magnolia in Handbuch der Laubgehölze, VOL.3, p.275-288. Meyer, F.G. - (1977) - Magnoliaceae in Flora of North America, VOL.3: online edition Rehder, A. - (1940) - Magnoliaceae in Manual of cultivated trees and shrubs hardy in North America, p.246-253. -

A Note on Magnolia, Mainly of Sections Manglietia and Michelia Subgenus
A note on Magnolia, mainly of sections Manglietia and Michelia subgenus Magnolia (Magnoliaceae) A note of caution concerning the ultimate heights that may be achieved in cultivation by numerous of the newer evergreen magnolias from Asia, is the theme of this article by CHRIS CALLAGHAN and S. K. PNG of the Australian Bicentennial Arboretum (ABA). Following Thomas Methuen-Campbell’s interesting report in the 2011 IDS Yearbook concerning the study weekend held in June of that year at RHS Wisley, Surrey, to discuss “summer” flowering magnolias (see Endnote), the authors thought they should write to mention an important consideration before contemplating planting of these trees in gardens, or indeed any tree in a garden, particularly the average small garden. We are not sure if the ultimate size of many of these magnolias was discussed with those attending the study weekend, since most of their maximum known heights were not mentioned in the article. However, we believe any readers tempted by the article to purchase and plant out most of the evergreen magnolias featured (previously in the genera Manglietia, Michelia or Parakmeria) in a normal suburban front or backyard in relatively 46 warm, sheltered, near frost-free areas, will be ultimately dismayed by the sizes they reach (see Figlar 2009 for reasons behind the reduction of genera). Even allowing that these predominantly warm-temperate to sub-tropical forest trees may not achieve their maximum potential sizes in the milder regions of temperate Europe, most are still likely to overtop (and overshadow!) two or three storey homes or apartments, especially with a warming climate. -

Phylogenomic Approach
Toward the ultimate phylogeny of Magnoliaceae: phylogenomic approach Sangtae Kim*1, Suhyeon Park1, and Jongsun Park2 1 Sungshin University, Korea 2 InfoBoss Co., Korea Mr. Carl Ferris Miller Founder of Chollipo Arboretum in Korea Chollipo Arboretum Famous for its magnolia collection 2020. Annual Meeting of Magnolia Society International Cholliop Arboretum in Korea. April 13th~22th, 2020 http://WWW.Chollipo.org Sungshin University, Seoul, Korea Dr. Hans Nooteboom Dr. Liu Yu-Hu Twenty-one years ago... in 1998 The 1st International Symposium on the Family Magnoliaceae, Gwangzhow Dr. Hiroshi Azuma Mr. Richard Figlar Dr. Hans Nooteboom Dr. Qing-wen Zeng Dr. Weibang Sun Handsome young boy Dr. Yong-kang Sima Dr. Yu-wu Law Presented ITS study on Magnoliaceae - never published Ten years ago... in 2009 Presented nine cp genome region study (9.2 kbp) on Magnoliaceae – published in 2013 2015 1st International Sympodium on Neotropical Magnoliaceae Gadalajara, 2019 3rd International Sympodium and Workshop on Neotropical Magnoliaceae Asterales Dipsacales Apiales Why magnolia study is Aquifoliales Campanulids (Euasterids II) Garryales Gentianales Laminales Solanales Lamiids important in botany? Ericales Asterids (Euasterids I) Cornales Sapindales Malvales Brassicales Malvids Fagales (Eurosids II) • As a member of early-diverging Cucurbitales Rosales Fabales Zygophyllales Celestrales Fabids (Eurosid I) angiosperms, reconstruction of the Oxalidales Malpighiales Vitales Geraniales Myrtales Rosids phylogeny of Magnoliaceae will Saxifragales Caryphyllales -
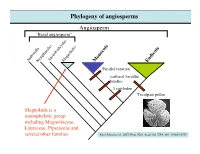
Phylogeny of Angiosperms Angiosperm “Basal Angiosperm”
Phylogeny of angiosperms Angiosperm “Basal angiosperm” AmborellaNymphaealesAustrobaileyalesMagnoliidss Monocots Eudicots Parallel venation scattered vascular bundles 1 cotyledon Tricolpate pollen Magnoliids is a monophyletic group including Magnoliaceae, Lauraceae, Piperaceae and several other families After Jansen et al., 2007, Proc. Natl. Acad. Sci. USA 104: 19369-19374 Magnoliaceae (Magnolia family) Textbook DVD KRR Magnolia X soulangeana; Magnoliaceae (Magnolia family) Textbook DVD WSJ Textbook DVD KRR Magnolia grandiflora; Magnolia macrophylla; Note leaf simple, entire, pinnate venation, numerous tepals, numerous stamens and carples. Textbook DVD KRR Magnolia sieboldii; Magnoliaceae (Magnolia family) Textbook DVD KMN Textbook DVD SMK-KRR Magnolia figo; Magnolia grandiflora; Note the elongated receptacle, Note the aggregate of follicles, and laminar stamens and red fleshy seed coat Magnoliaceae (Magnolia family) Photo: Yaowu Yuan Photo: Yaowu Yuan Liriodendron tulipifera; Note the elongated receptacle, and laminar stamens Magnoliaceae (Magnolia family) Note the lobed, T-shirt-like leaf, and pinnate venation Note the aggregate of samara Magnoliaceae (Magnolia family) Magnoliaceae - 2 genera/220 species. Trees or shrubs; Ethereal oils (aromatic terpenoids) - (remember the smell of bay leaves?); Leaves alternate, simple (Magnolia) or lobed (Liriodendron), entire; Flowers large and showy, actinomorphic, bisexual Tepals 6-numerous, stamens and carpels numerous, Spirally arranged on an elongated receptacle, Laminar stamens poorly differentiated into anther and filament. Fruit usually an aggregate of follicle (Magnolia) or samara (Liriodendron); follicle: 1-carpellate fruit that dehisces on the side samara: 1-carpellate winged, indehiscent fruit Phylogeny of Eudicots (or Tricolpates) Eudicots (or Tricolpates) “Basal eudicots” Asterids Buxales Rosids Caryophyllales RanunculalesProteales Ranunculales is a monophyletic group including Ranunculaceae, Berberidaceae, Papaveraceae, and 4 other families. After Jansen et al., 2007, Proc. -

Vascular Plant Families of the United States Grouped by Diagnostic Features
Humboldt State University Digital Commons @ Humboldt State University Botanical Studies Open Educational Resources and Data 12-6-2019 Vascular Plant Families of the United States Grouped by Diagnostic Features James P. Smith Jr Humboldt State University, [email protected] Follow this and additional works at: https://digitalcommons.humboldt.edu/botany_jps Part of the Botany Commons Recommended Citation Smith, James P. Jr, "Vascular Plant Families of the United States Grouped by Diagnostic Features" (2019). Botanical Studies. 96. https://digitalcommons.humboldt.edu/botany_jps/96 This Flora of the United States and North America is brought to you for free and open access by the Open Educational Resources and Data at Digital Commons @ Humboldt State University. It has been accepted for inclusion in Botanical Studies by an authorized administrator of Digital Commons @ Humboldt State University. For more information, please contact [email protected]. FLOWERING PLANT FAMILIES OF THE UNITED STATES GROUPED BY DIAGNOSTIC FEATURES James P. Smith, Jr. Professor Emeritus of Botany Department of Biological Sciences Humboldt State University Second edition — 6 December 2019 The focus is on families of plants found in the conterminous United States, including ornamentals. The listing of a family is not meant to imply that every species has that feature. I am using a fewfamily names, such as Liliaceae, Plantaginaceae, and Scrophulariaceae, in the traditional sense, because their limits remain unsettled. Parasitic on branches Dioscoreaceae -

Effective Heat Transport During Miocene Cooling
EGU Journal Logos (RGB) Open Access Open Access Open Access Advances in Annales Nonlinear Processes Geosciences Geophysicae in Geophysics Open Access Open Access Natural Hazards Natural Hazards and Earth System and Earth System Sciences Sciences Discussions Open Access Open Access Atmospheric Atmospheric Chemistry Chemistry and Physics and Physics Discussions Open Access Open Access Atmospheric Atmospheric Measurement Measurement Techniques Techniques Discussions Discussion Paper | Discussion Paper | Discussion Paper | Discussion Paper | Open Access Biogeosciences Discuss., 10, 13563–13601, 2013 Open Access www.biogeosciences-discuss.net/10/13563/2013/ Biogeosciences Biogeosciences BGD doi:10.5194/bgd-10-13563-2013 Discussions © Author(s) 2013. CC Attribution 3.0 License. 10, 13563–13601, 2013 Open Access Open Access This discussion paper is/has been under review for the journal BiogeosciencesClimate (BG). Climate Effective heat Please refer to the correspondingof finalthe Past paper in BG if available. of the Past Discussions transport during Miocene cooling Open Access Open Access Earth System Earth System T. Denk et al. Effective heat transportDynamics of Gulf StreamDynamics to Discussions subarctic North Atlantic during Miocene Title Page Open Access Geoscientific Geoscientific Open Access cooling: evidenceInstrumentation from “KöppenInstrumentation Abstract Introduction Methods and Methods and signatures” ofData fossil Systems plant assemblagesData Systems Conclusions References Discussions Open Access Open Access Tables Figures 1 1 2 2 Geoscientific T. Denk , G. W. Grimm , F.Geoscientific Grímsson , and R. Zetter Model Development 1 Model Development J I Swedish Museum of Natural History, Department of Palaeobotany, Box 50007,Discussions 10405 Stockholm, Sweden 2 J I Open Access University of Vienna, Department of Palaeontology,Open Access Althanstrasse 14, 1090 Vienna, Austria Hydrology and Hydrology and Back Close Received: 8 July 2013 – Accepted:Earth System 31 July 2013 – Published: 15 AugustEarth 2013 System Correspondence to: T. -

Pignut Hickory
Carya glabra (Mill.) Sweet Pignut Hickory Juglandaceae Walnut family Glendon W. Smalley Pignut hickory (Curya glabru) is a common but not -22” F) have been recorded within the range. The abundant species in the oak-hickory forest associa- growing season varies by latitude and elevation from tion in Eastern United States. Other common names 140 to 300 days. are pignut, sweet pignut, coast pignut hickory, Mean annual relative humidity ranges from 70 to smoothbark hickory, swamp hickory, and broom hick- 80 percent with small monthly differences; daytime ory. The pear-shaped nut ripens in September and relative humidity often falls below 50 percent while October and is an important part of the diet of many nighttime humidity approaches 100 percent. wild animals. The wood is used for a variety of Mean annual hours of sunshine range from 2,200 products, including fuel for home heating. to 3,000. Average January sunshine varies from 100 to 200 hours, and July sunshine from 260 to 340 Habitat hours. Mean daily solar radiation ranges from 12.57 to 18.86 million J mf (300 to 450 langleys). In Native Range January daily radiation varies from 6.28 to 12.57 million J m+ (150 to 300 langleys), and in July from The range of pignut hickory (fig. 1) covers nearly 20.95 to 23.04 million J ti (500 to 550 langleys). all of eastern United States (11). It extends from According to one classification of climate (20), the Massachusetts and the southwest corner of New range of pignut hickory south of the Ohio River, ex- Hampshire westward through southern Vermont and cept for a small area in Florida, is designated as extreme southern Ontario to central Lower Michigan humid, mesothermal. -

Assessing the Population Genetic Structure of the Endangered Cucumber Tree (Magnolia Acuminata) in Southwestern Ontario Using Nuclear and Chloroplast Genetic Markers
ASSESSING THE POPULATION GENETIC STRUCTURE OF THE ENDANGERED CUCUMBER TREE (MAGNOLIA ACUMINATA) IN SOUTHWESTERN ONTARIO USING NUCLEAR AND CHLOROPLAST GENETIC MARKERS. A Thesis Submitted to the Committee on Graduate Studies in Partial Fulfillment to the Requirements for the Degree of Master of Science in the Faculty of Arts and Science Trent University Peterborough, Ontario, Canada © Copyright by Cara E. Budd 2014 Environmental and Life Sciences M.Sc. Graduate Program September 2014 Assessing the population genetic structure of the endangered Cucumber tree (Magnolia acuminata) in southwestern Ontario using nuclear and chloroplast genetic markers. Cara E. Budd ABSTRACT Magnolia acuminata (Cucumber tree) is the only native Magnolia in Canada, where it is both federally and provincially listed as endangered. Magnolia acuminata in Canada can be found inhabiting pockets of Carolinian forest within Norfolk and Niagara regions of southwestern Ontario. Using a combination of nuclear and chloroplast markers, this study assessed the genetic diversity and differentiation of M. acuminata in Canada, compared to samples from the core distribution of this species across the United States. Analyses revealed evidence of barriers to dispersal and gene flow among Ontario populations, although genetic diversity remains high and is in fact comparable to levels of diversity estimated across the much broader range of M. acuminata in the USA. When examining temporal differences in genetic diversity, our study found that seedlings were far fewer than mature trees in Ontario, and in one site in particular, diversity was lower in seedlings than that of the adult trees. This study raises concern regarding the future viability of M. acuminata in Ontario, and conservation managers should factor in the need to maintain genetic diversity in young trees for the long-term sustainability of M. -

Magnolia Champaca and Magnolia × Alba
Comparison of Similar Looking Plants Magnolia champaca and Magnolia × alba © Zaki Jamil © Zaki Jamil Scientific name: Magnolia champaca Scientific name: Magnolia × alba Common name: Champaca Common name: White Champaca Family: Magnoliaceae Family: Magnoliaceae Origin: India Origin: Cultivated hybrid © Horticulture Outreach and Heritage Trees, National Parks Board, 2019 Character Comparison Magnolia champaca Magnolia × alba Form • Up to 40-50 m tall • Up to 10-30 m tall • Conical, compact crown • More open crown Foliage • Spirally arranged, ovate- • Spirally arranged, lanceolate to oblong- ovate-lanceolate to lanceolate leaves (10-30 oblong-lanceolate cm long, 4-10 cm wide) leaves (15-35 cm • Leaf stalk 1-4 cm long long, 5.5-16 cm wide) • Leaf stalk 1.5-5 cm long © Horticulture Outreach and Heritage Trees, National Parks Board, 2019 Character Comparison Magnolia champaca Magnolia × alba Foliage • Matte leaves with velvety • Glossy leaves with smooth Comparison underside (hairs more underside dense along the midrib and veins) Stipule- • Long stipule-scar almost as • Short stipule-scar located Scar long as the leaf stalk at the base of the leaf stalk © Cerlin Ng © Horticulture Outreach and Heritage Trees, National Parks Board, 2019 Character Comparison Magnolia champaca Magnolia × alba Flower • Solitary flowers (4-5 cm • Solitary flowers (5 cm wide) have an intense, wide) have a more subtle, musky fragrance fruity fragrance • Flowers light yellow • Flowers are white (no turning to dark orange colour change) (flowers of some wild • Free-flowering, abundant plants may remain light blooming yellow) • Occasional flowering © Cerlin Ng Fruit • Each flower produces a • Sterile hybrid does not cluster of round to egg- produce fruit shaped, light brown, slightly woody fruit (called a ‘follicle’, 1.5-3.5 cm long, 1-2.5 cm wide) © Horticulture Outreach and Heritage Trees, National Parks Board, 2019 Character Comparison References: Gardner, S., Sidisunthorn, P.