Deer Density and Plant Palatability Predict Shrub Cover, Richness
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

51 Salmon. Returning Through the San Juan and Gulf Islands
salmon. Returning through the San Juan and Gulf Islands throughout August and September, the cycle ended with chum salmon as the last major resource harvested in the Goldstream area from October onward. (2) Image 1: The W̱ SÁNEĆ Calendar, artist Briony Penn, copyright Tsawout First Nation. W̱ SÁNEĆ Elders Earl Claxton and John Elliot created a written version of the tradit ional 13-moon W̱SÁNEĆ calendar in 1993, which is included below: 51 The W̱SÁNEĆ Year ṈIṈENE – Moon of the Child ♦ Nettle twine made into nets. ♦ For those who would take the risk, Halibut fishing resumed. ♦ Grilse fishing in WJOLEP. ♦ Fawns born, deer hunting stops. ♦ Generally living off supplies, telling tales, instructing children, longhouse ceremonies continue. WEXES – Moon of the Frog ♦ Ocean travel safer. ♦ Fish for herring and gather herring roe. ♦ Duck nets were used. ♦ Sections of reef net sewn together, captains surveyed beaches for anchor rocks, rafts were assembled, floats and cables prepared. PEXSISEṈ – The Moon of Opening Hands – The Blossoming Out Moon ♦ Hunting of Brant Geese ♦ Clams, oysters and Mussels are at their best. ♦ Cedar logs felled for canoes, etc. ♦ Cedar bark gathered. ♦ Wool dogs shed – wool gathered. SXÁNEȽ - Bullhead Moon ♦ Gathered Bullheads. ♦ Harvested seaweed. ♦ Halibut fishery ends. ♦ Grouse snared in the woods. PENÁW̱EṈ – Moon of Camas Harvest ♦ Most families left the winter village for spring camps. ♦ Harvest camas. ♦ Gather gull eggs. ♦ Purple and green sea urchins gathered. ĆENŦEḴI – The Sockeye Moon ♦ Dropping of the anchor rocks at the family reef net locations. ♦ First salmon ceremony ♦ Start of salmon trade. ĆENHENEN – Humpback Salmon Return to the Earth 52 ♦ Reef fishery continues – especially at Point Roberts. -

Bald Eagles Wintering in the Southern Gulf Islands, British Columbia
BALD EAGLES WINTERING IN THE SOUTHERN GULF ISLANDS, BRITISH COLUMBIA DAVID HANCOCK HIS study represents the first attempt to census regularly the wintering T Bald Eagles Haliaeetus Zeucocephdus in the southern Gulf Islands, Brit- ish Columbia. Data were also collected on the feeding and hunting habits of the wintering eagles. In 1961 the National Audubon Society initiated its 5-year Continental Bald Eagle Study. This North American survey was prompted largely by the alarming decrease in numbers of the eastern Bald Eagle in recent years. The studies of Broley (1947)) Howell (1962)) and others have amply pointed out the plight of this great bird in the eastern United States. More recently Southern (1963) conducted a survey of the Bald Eagles winter- ing along the Mississippi River in northwestern Illinois. No similar studies have been conducted on the west coast prior to the Continental Bald Eagle project, or to my study. Murie (1940 and 1959) described the food habits of the eagles of the Aleutian Islands, and Dixon (1909) gives a brief history of the Alaskan birds. Brooks (1922) and Munro (1938) p resented a few of their observations on the feeding and hunting habits of Bald Eagles in British Columbia. Less important notes on individual sightings, nestings, and feeding incidents complete the literature on this species on the west coast. METHOD Fifteen aerial counts, involving 24 hours and 15 minutes of flying time, were conducted over the area (Fig. 1) from 26 September 1962 to IS April 1963. In addition, 130 hours were spent on ground observations. The study area (shown in Fig. -

Bedrock Geology of the North Saanich-Cobble Hill Area, British Columbia, Canada
AN ABSTRACT OF THE THESIS OF John Michael Kachelmeyer for the degree ofMaster of Science in Geology presented on May 30, 1978 Title: BEDROCK GEOLOGY OF THE NORTHSAANICH-COBBLE HILL AREAS, BRITISH COLUMBIA, CANADA Abstract approved: Redacted for Privacy Keith F./Oles The bedrock of the North Saanich-Cobble Hill areas consists of igneous and sedimentary rocks of Early Jurassic through Late Cretaceous age.Early Jurassic andesitic Bonanza Volcanics are intruded by Middle Jurassic Saanich Granodiorite plutonic rocksand dikes.Late Cretaceous Nanaimo Group clastic rocksnonconformably overlie these Jurassic units.The four oldest formations of the group (Comox, Has lam, Extension-Protection, and Cedar District) are exposed within the study area. The Nanaimo Group formations were deposited within a subsid- ing marine basin (the Nanaimo Basin) located to the east ofsouthern Vancouver Island.Uplift of pre-Late Cretaceous basement rocks on Vancouver Island to the west and on mainland British Columbia tothe east of the basin created rugged, high-relief source areasthat were chemically weathered in a warm tropical climate and mechanically eroded by high-energy braided stream systems. The Comox and Has lam sediments were deposited in response to a continual transgression of a Late Cretaceous seaway overthis rugged terrain.Braided stream systems trending north-northwest into the basin deposited Comox (Benson Member) gravelsand sands onto a high-energy, cliffed shoreline.Erosion of the highlands and transgression of the marine seaway resulted in subduedtopographic relief and the deposition of sediments by low-energymeandering streams within a river traversed coastal plain, followedby silt and clay deposition in tidal flat and lagoonal environments,and finally by sand deposition in a higher energy barrier bar environment.Contin- ued marine transgression resulted in the depositionof massive marine mudstones and cyclic sandstone to mudstone sequencesof the Has lam Formation. -
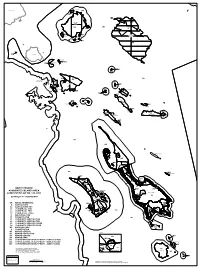
North Pender Associated Islands Area Land Use Bylaw No. 148, 2003
North Pender I. Chads Island Salt Spring Island L 38 W4 RR ´ 9 Portland RR Island 8 Pellow Islets NP W1 NP A 7 6 NP RR Brackman Moresby Island Tortoise Islets Island R 1W R 1E RR 5 Hood Island NP RR 4 3 Arbutus I. 2 RR Piers Island 1 Pl 20349 Pym Island L 103 A RR(a) Knapp Island RR L 102 Imrie Island NP W4 W3 RR W1 Swartz Bay Ferry Terminal RR W4 Musclow I. Goudge L 94 Island I1 Coal Island L 95 NP L 93 RR A Reay Island Johnson I. 37 L 96 Fernie Island W4 Brethour Island RR RR A L 97 Greig Island NP Sheep RR Island RL 1R08 C A U N Domville S A Dock A RR NP Island D Island A Little Shell W4 RR Island L 91 L 98 Ker Island L 101 RR Comet Island RR Rubly L 107 Island W4 CD1 RR L 100 Gooch Island Forrest Island NP Rum Island L 99 RR L 71 Sidney W1 W4 NP Mandarte Island 940R/W NP 1 NP 402R CD2(a) 122 123 19 B 121 CD2 124 y la W2 125 w 119 W2 E RR x L 129 Halibut Island te n 117 t B 126 o u 115 127 n W1 d a r y 113 2 3 4 W3 1 3 1 131 6 111 128 3 1 27 129 S1 0 G 3 CD3(a) 1 109 CD2 29 31 4 W2 107 G S1 33 CD3 G 35 W2 37 105 G 39 41 NORTH PENDER 103 CD2 CD2 43 VIS 5122 45 S4 SEE INSET ASSOCIATED ISLANDS AREA 47 101 G S3 Sidney Island 49 LAND USE BYLAW NO. -

Gulf Islands
Education Young at Week Heart SD 64 schools & INSIDE A nnivers arary programs. B section 1960-2010 GULF ISLANDS Wednesday, March 17, 2010 — YOUR COMMUNITY NEWSPAPER SINCE 1960 50TH YEAR ISSUE 11 $ 25 1(incl. GST) COURT ACHIEVEMENTS Island man Province nnivers ary recognizes enters guilty Murakami generosity plea in Richard Murakami one of 45 B.C. Community knife attack Achievement Award winners Kitchen knife used in throat BY SEAN MCINTYRE DRIFTWOOD STAFF slashing At this rate, the province’s Lieutenant Governor should BY SEAN MCINTYRE just hand over the house DRIFTWOOD STAFF keys. “You have to come over, something terrible has Less than a year after Salt happened,” Simon Teskey told his mother in a Spring’s Richard Murakami telephone call only moments after a March 2009 visited Victoria’s Govern- drug deal took a turn for the worse. ment House to meet the Teskey was in Ganges Provincial Court last Emperor and Empress of week facing charges of attempted murder and Japan, he’s been asked to aggravated assault stemming from an incident in come down and make yet which Crown counsel said he used a “ridiculously another appearance. big French chef’s knife” to slash the neck of an This time around, acquaintance who had come to collect a $200 drug Murakami and 44 other Brit- debt. ish Columbia Community Court heard that Teskey, 30, had been partying Achievement Award win- off and on with the victim and three other island ners will be celebrated as the residents since the previous day when the early guests of honour during an afternoon incident took place at Teskey’s Bull- April 28 gala. -

SOUTHERN GULF ISLANDS HARBOURS COMMISSION Notice of a Meeting on Friday, August 25, 2017 at 9 Am Saanich Peninsula Health Unit, 2170 Mt
SOUTHERN GULF ISLANDS HARBOURS COMMISSION Notice of a meeting on Friday, August 25, 2017 at 9 am Saanich Peninsula Health Unit, 2170 Mt. Newton X Road, Saanichton, BC D. Hargreaves, N. Pender Island L. Peck, Saturna Island D. Maude, Mayne Island R. Fenton, S. Pender Island B. Young, Piers Island D. Howe, SGI EA Director B. Mabberley, Galiano Island AGENDA 1. Approval of the Agenda 2. Adoption of the Minutes of the Meeting held June 9, 2017 3. Chair’s Remarks 4. Presentations/Delegations • Brian Wheeler, regarding the CRD Dock on Piers Island 5. Correspondence (a) Dan MacDonald, Seair Seaplanes, July 12, 2017, regarding Mayne Island Seaplane Dock Relocation (b) Brian Wheeler, July 16, 2017, regarding Piers Island Dock 6. Miners Bay Dock – CRD/BC Ferries Update (Verbal) 7. Operating Budget Update - Attached 8. Dock Operations Reports – Attached 9. 2017 Capital Plan Work Schedule – Attached 10. Lyall Harbour Dock Divestiture Update (Verbal) 11. New Business 12. Adjournment To ensure a quorum, please advise Margaret at 250.474.9606 if you or your alternate cannot attend. Minutes of a Meeting of the Southern Gulf Islands Harbour Commission Held June 9, 2017 at the Saanich Peninsula Health Unit, Mt. Newton X Road, Victoria, BC PRESENT: COMMISSIONERS: B. Mabberley, Galiano Island; David Maude, Mayne Island; B. Young, Piers Island; L. Peck, Saturna Island; Dave Hargreaves, N Pender Island; D. Howe, Regional Director. STAFF: Ted Robbins, General Manager, Integrated Water Services; Dale Puskas, Manager, Capital Works; Dan Robson, Manager, Saanich Peninsula and Gulf Islands Operations; S. Orr (recorder); Peter Binner, Dock Operations Coordinator ABSENT: R. -

SOUTHERN GULF ISLANDS HARBOURS COMMISSION Notice of a Meeting on Friday, January 27, 2017 at 9 Am Saanich Peninsula Health Unit, 2170 Mt
SOUTHERN GULF ISLANDS HARBOURS COMMISSION Notice of a meeting on Friday, January 27, 2017 at 9 am Saanich Peninsula Health Unit, 2170 Mt. Newton X Road, Saanichton, BC D. Hargreaves, N. Pender Island L. Peck, Saturna Island D. Maude, Mayne Island R. Fenton, S. Pender Island B. Young, Piers Island D. Howe, SGI EA Director B. Mabberley, Galiano Island d AGENDA 1. Approval of the Agenda e 2. Adoption of the Minutes of the Meeting held October 28, 2016 ........................................l 1 3. Chair’s Remarks u 4. Presentations/Delegations • No one has registered to speak. d 5. Miners Bay Dock Facility – Vessel Wake Assessment and Conceptual Dock Layout (Report # SGIHC 2017-01) ..............................................................................................e 5 6. Southern Gulf Islands Harbours Service – Amended 2017 Operating and Capital Budget (Report #SGIHC 2017-02) .............................................................................................h 57 7. Piers Island Dock Facility - Floatc Geometry Review (Report #SGIHC 2017-03) ............. 88 8. Dock Capital Upgrade sProject Status Update 9. Status of Wharfinger Service Agreements (Verbal Report) 10. Commissioner eand Wharfinger Contact Information..................................................... 113 11. Status Rof Dock Operations Coordinator Services Agreement (Verbal Report) 12. Adjournment To ensure a quorum, please advise Margaret at 250.474.9606 if you or your alternate cannot attend. 1 1 Minutes of a Meeting of the Southern Gulf Islands Harbour Commission Held October 28, 2016 at the Saanich Peninsula Health Unit, Mt. Newton X Road, Victoria, BC PRESENT: Commission: B. Mabberley, Galiano Island; Carl Bunnin, Mayne Island B. Young, Piers Is.; D. Howe, Regional Director; Paul LeBlond, Alternate Director Staff: Ted Robbins, General Manager, Integrated Water Services; Peggy Dayton, Sr. Financial Analyst; Ian Sander, Capital Projects Manager; Dan Robson, Manager, Saanich Peninsula and Gulf Islands Operations; M. -

Hawaiian Settlement in British Columbia
JEAN BARMAN New Land, New Lives: Hawaiian Settlement in British Columbia BETWEEN THE LATE eighteenth and mid-nineteenth centuries a thou- sand or more, perhaps several thousand, native Hawaiians visited the west coast of North America as seamen, fur trade laborers, or independent adventurers.1 While most returned home, a handful stayed behind. Patterns of Hawaiian settlement and family formation in British Columbia are distinctive from those of Hawaiians elsewhere in the Pacific Northwest. Immediately to the south, Hawaiians were subjected to the legalized discrimination inflicted on Indians and blacks, whereas those who settled in British Columbia were accorded the same civil rights enjoyed by members of the dominant society. The consequence has been that, despite some racism based on physi- cal appearance and aboriginal origin, descendants have not become segregated, or segregated themselves, but rather married and lived in ways similar to others of comparable socioeconomic status. Hawaiian men began to arrive on the west coast of North America almost from the time British sea captain James Cook visited the Hawaiian Islands, named by him the Sandwich Islands, in January 1778.2 The islands quickly became a wintering and stopover point for European and American merchant vessels. Local men—sometimes termed Owyhees, an adaptation of Hawaiians, but more often called Kanakas, a Polynesian term meaning simply a human being or per- son Barman is a professor in the Department of Educational Studies at the University of British Columbia. She is the author of The West beyond the West: A History of British Columbia (1991, 2nd ed. 1996) and co-editor of the journalBC Studies. -

Islands Trust - Local Trustees
Islands Trust - Local Trustees Notice of Election by Voting | 2014 General Local Election Public notice is hereby given that General Voting Day will be held from 8:00 am to 8:00 pm Mail Ballot Voting is now available to electors who: on Saturday, November 15, 2014 to elect persons to serve a four-year term of office from • have a physical disability, illness or injury that affects their ability to vote at another voting December 2014 until after the 2018 election held in accordance with the legislation in the Local opportunity, or Government Act) as Local Trustees for the six island trust areas under the Islands Trust. • expect to be absent from the regional district on general voting day (Saturday, November 15, 2014) Galiano Island Local Trust Area - Two Trustees to be Elected: and at the times of all advance voting opportunities (Wednesday, November 5 and 12, 2014). Surname Given Name(s) Residential Address You must be a qualified Resident or Non-Resident property elector to vote. AGES Dave 391 Cain Road, Galiano Island, BC To Register To Vote By Mail, eligible electors who meet the criteria must submit a completed HARRIS George Robert 520 Cameron Lane, Galiano Island, BC application form in advance of general voting day, November 15, 2014. Application forms are POTTLE Sandy 5630 Porlier Pass Road, Galiano Island, BC available on the CRD website at www.crd.bc.ca/mailballot. Completed application forms may be Mayne Island Local Trust Area - Two Trustees to be Elected: submitted via email, mail, in person or via fax at the following contact information listed below: Surname Given Name(s) Residential Address Capital Regional District, Legislative & Information Services CRUMBLEHULME Brian 639 Horton Bay Road, Mayne Island, BC 5th floor, 625 Fisgard Street, PO Box 1000, Victoria, BC, V8W 2S6 DODDS Jeanine Gay 720 Simpson Road, Mayne Island, BC Phone: 250.360.3129 | Toll free: 1.800.663.4425 local 3129 MAUDE David P. -

2019-06-28 Pauquachin First Nation Response to RBT2 Review Panel
Response to Information Request from Roberts Bank Terminal 2 Project Review Panel at the Public Hearing Session on June 20, 2019 Prepared by Dr. Peter Evans (Trailmark Systems) and submitted on behalf of Pauquachin First Nation Methods The results analysis presented to the Proponent and to the Panel on June 20, 2019, and expanded on below, drew on several different types of information, including ethnohistorical research and analysis, ethnography, map biography interviews, participatory group mapping, AIS data from the Canadian Coast Guard, GIS analysis, and previous research. The traditional use information in the W’SANEC Traditional Use Study (CEAR #1670) came from five different studies: the Sencoten Alliance Study (2002); the Parks Canada Study (2013); the TMX Study (2013); and the Places of the W’SANEC Nation Study (Fritz 2016). Three of the four projects were community-based, while one (Fritz 2016) was led by a university graduate researcher. Each used a slightly different methodology. Sencoten Alliance (2002) relied on the Tobias method (Tobias 2000; 2009); the Parks Canada study (2013) an Indigenous research method; the TMX Study a mixed method of qualitative oral history interviews and participatory mapping; and the Places of the W’SANEC Nation study used an ethnohistorical mapping method that relied mostly on texts and participatory engagement. Variation on the “map biography” method formed the main technique used in three of the four traditional land and marine use studies. This method emerged as the dominant research technique for documenting land use extensity and occupation after the rise in Indigenous land claims in the 1970s. Its popularity is due in part to its centring of GIS as an appeal to scientific objectivity. -

To Ensure Quorum, Advise Denise Dionne, 250-360-3087 If You Cannot Attend
1 1 SOUTHERN GULF ISLANDS HARBOURS COMMISSION Notice of Meeting on Friday, June 25, 2021 at 9:30 a.m. Goldstream Meeting room, 479 Island Highway, Victoria, BC For members of the public who wish to listen to the meeting via telephone please call 1-833-353-8610 and enter the Participant Code 1911461 followed by #. You will not be heard in the meeting room but will be able to listen to the proceedings. B. Dearden (Chair), Mayne Island J. Hall, Piers Island/Swartz Bay P. Brent (V. Chair), Saturna Island D. Howe, Electoral Area Director J. Deschenes, North Pender Island B. Mabberley, Galiano Island R. Fenton, South Pender Island AGENDA 1. APPROVAL OF AGENDA 2. ADOPTION OF MINUTES ....................................................................................................3 Recommendation: That the minutes of the April 23, 2021 meeting be adopted. 3. CHAIR’S REMARKS 4. PRESENTATIONS/DELEGATIONS In keeping with directives from the Province of BC, this meeting will be held without the public present. A phone in number is provided above that will allow the public to listen to the meeting. Presentation and Delegation requests can be made online or complete this printable form (PDF). Requests must be received no later than 4:30 p.m. two calendar days prior to the meeting. 5. COMMISSION BUSINESS 5.1. Project Update – June 2021 .................................................................................... 10 Recommendation: That the Southern Gulf Islands Harbours Commission receive this report for information. 5.2. West Coast Marine Response Corporation (WCMRC) Update [Verbal] 5.3. BC Ferries – Sturdies Bay Update .......................................................................... 12 Recommendation: That the update from BC Ferries be received for information. To ensure quorum, advise Denise Dionne, 250-360-3087 if you cannot attend. -

Gulf Islands
All wet Margo abroad $ 25 Folks take the plunge at Salt Spring youth talks about 1(incl. GST) Fulford and Vesuvius. experience in Ghana. PAGE 2 PAGE 4 GULF ISLANDS Wednesday, January 6, 2010 — YOUR COMMUNITY NEWSPAPER SINCE 1960 50TH YEAR ISSUE 1 FIRE Quick action stops fi re in its tracks Ganges pasta shop re-opens today BY GAIL SJUBERG DRIFTWOOD EDITOR Storing paper products next to a pot light in a closet caused a minor fi re at Pasta Fresca del Vecchio last Thursday morn- ing. Salt Spring fi refi ghters were called to the Gasoline Alley/ Restaurant Row establishment at 8:26 a.m., where they quickly extinguished the flames and removed smouldering materi- als from the building. Business co-owner Carly Goring said it was fortunate PHOTO BY DERRICK LUNDY the fi re was stopped before it could spread throughout the ROCKING NEW YEAR’S: Harry Manx is the man of the midnight hour as he performs to a full house at Fulford Hall on New Year’s Eve. The event building that houses several served as a benefi t for the Wolf Kids educational program on Salt Spring Island. businesses. “We’re really lucky I was PROPERTY ASSESSMENTS down here when it started,” she said. With a fi re extinguisher she was able to prevent the fire in the ceiling from growing Gulf Islands land values take a dip beyond the kitchen area. In addition to the impact of Trend diff ers from province-wide stats that saw an Gulf Islands property owners can expect to the extinguisher chemicals, see their notices in the mail any day if they Salt Spring Fire Chief Tom overall increase in property values haven’t already received them.