Antidepressant Drug-Induced Hypothalamic Cooling in Syrian Hamsters Wallace C
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

HALDOL Brand of Haloperidol Injection (For Immediate Release) WARNING Increased Mortality in Elderly Patients with Dementia
HALDOL® brand of haloperidol injection (For Immediate Release) WARNING Increased Mortality in Elderly Patients with Dementia-Related Psychosis Elderly patients with dementia-related psychosis treated with antipsychotic drugs are at an increased risk of death. Analyses of seventeen placebo-controlled trials (modal duration of 10 weeks), largely in patients taking atypical antipsychotic drugs, revealed a risk of death in drug-treated patients of between 1.6 to 1.7 times the risk of death in placebo-treated patients. Over the course of a typical 10-week controlled trial, the rate of death in drug-treated patients was about 4.5%, compared to a rate of about 2.6% in the placebo group. Although the causes of death were varied, most of the deaths appeared to be either cardiovascular (e.g., heart failure, sudden death) or infectious (e.g., pneumonia) in nature. Observational studies suggest that, similar to atypical antipsychotic drugs, treatment with conventional antipsychotic drugs may increase mortality. The extent to which the findings of increased mortality in observational studies may be attributed to the antipsychotic drug as opposed to some characteristic(s) of the patients is not clear. HALDOL Injection is not approved for the treatment of patients with dementia-related psychosis (see WARNINGS). DESCRIPTION Haloperidol is the first of the butyrophenone series of major antipsychotics. The chemical designation is 4-[4-(p-chlorophenyl)-4-hydroxypiperidino] 4’-fluorobutyrophenone and it has the following structural formula: HALDOL (haloperidol) is available as a sterile parenteral form for intramuscular injection. The injection provides 5 mg haloperidol (as the lactate) and lactic acid for pH adjustment between 3.0 – 3.6. -

Dosulepin Prescribing
SAFETY BULLETIN: DOSULEPIN PRESCRIBING In December 2007, the Medicines and Healthcare Regulatory Agency (MHRA) issued safety advice around prescribing of dosulepin, related to the narrow margin between therapeutic doses and potentially fatal doses. Nevertheless, dosulepin continues to be prescribed widely. Over eight years after the safety advice was published, prescribing of dosulepin in the Dorset area remains high. In the South West area, Dorset CCG was the highest prescriber of dosulepin for the 2015/16 financial year (3.238% of all antidepressant items). Of the 209 CCGs in the UK, Dorset is the fourteenth highest prescriber of dosulepin, and well above the national average of 2.111%. The following points summarise the reasons that dosulepin is not recommended for prescribing nationally, and in Dorset: Dosulepin has a small margin of safety between the (maximum) therapeutic dose and potentially fatal doses. The NICE guideline on depression in adults recommends that dosulepin should not be prescribed for adults with depression because evidence supporting its tolerability relative to other antidepressants is outweighed by the increased cardiac risk and toxicity in overdose. Dosulepin has also been used ‘off label’ in other indications such as fibromyalgia and neuropathic pain. However the evidence for use in in this way is weak, and is not recommended. The lethal dose of dosulepin is relatively low and can be potentiated by alcohol and other CNS depressants. Dosulepin overdose is associated with high mortality and can occur rapidly, even before hospital treatment can be received. Onset of toxicity occurs within 4-6 hours. Every year, up to 200 people in England and Wales fatally overdose with dosulepin. -

Clomipramine | Memorial Sloan Kettering Cancer Center
PATIENT & CAREGIVER EDUCATION Clomipramine This information from Lexicomp® explains what you need to know about this medication, including what it’s used for, how to take it, its side effects, and when to call your healthcare provider. Brand Names: US Anafranil Brand Names: Canada Anafranil; MED ClomiPRAMINE; TARO-Clomipramine Warning Drugs like this one have raised the chance of suicidal thoughts or actions in children and young adults. The risk may be greater in people who have had these thoughts or actions in the past. All people who take this drug need to be watched closely. Call the doctor right away if signs like low mood (depression), nervousness, restlessness, grouchiness, panic attacks, or changes in mood or actions are new or worse. Call the doctor right away if any thoughts or actions of suicide occur. This drug is not approved for use in all children. Talk with the doctor to be sure that this drug is right for your child. What is this drug used for? It is used to treat obsessive-compulsive problems. It may be given to you for other reasons. Talk with the doctor. Clomipramine 1/8 What do I need to tell my doctor BEFORE I take this drug? If you have an allergy to clomipramine or any other part of this drug. If you are allergic to this drug; any part of this drug; or any other drugs, foods, or substances. Tell your doctor about the allergy and what signs you had. If you have had a recent heart attack. If you are taking any of these drugs: Linezolid or methylene blue. -
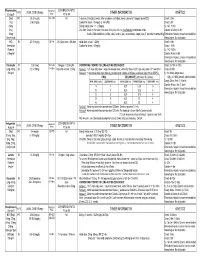
Medication Conversion Chart
Fluphenazine FREQUENCY CONVERSION RATIO ROUTE USUAL DOSE (Range) (Range) OTHER INFORMATION KINETICS Prolixin® PO to IM Oral PO 2.5-20 mg/dy QD - QID NA ↑ dose by 2.5mg/dy Q week. After symptoms controlled, slowly ↓ dose to 1-5mg/dy (dosed QD) Onset: ≤ 1hr 1mg (2-60 mg/dy) Caution for doses > 20mg/dy (↑ risk EPS) Cmax: 0.5hr 2.5mg Elderly: Initial dose = 1 - 2.5mg/dy t½: 14.7-15.3hr 5mg Oral Soln: Dilute in 2oz water, tomato or fruit juice, milk, or uncaffeinated carbonated drinks Duration of Action: 6-8hr 10mg Avoid caffeinated drinks (coffee, cola), tannics (tea), or pectinates (apple juice) 2° possible incompatibilityElimination: Hepatic to inactive metabolites 5mg/ml soln Hemodialysis: Not dialyzable HCl IM 2.5-10 mg/dy Q6-8 hr 1/3-1/2 po dose = IM dose Initial dose (usual): 1.25mg Onset: ≤ 1hr Immediate Caution for doses > 10mg/dy Cmax: 1.5-2hr Release t½: 14.7-15.3hr 2.5mg/ml Duration Action: 6-8hr Elimination: Hepatic to inactive metabolites Hemodialysis: Not dialyzable Decanoate IM 12.5-50mg Q2-3 wks 10mg po = 12.5mg IM CONVERTING FROM PO TO LONG-ACTING DECANOATE: Onset: 24-72hr (4-72hr) Long-Acting SC (12.5-100mg) (1-4 wks) Round to nearest 12.5mg Method 1: 1.25 X po daily dose = equiv decanoate dose; admin Q2-3wks. Cont ½ po daily dose X 1st few mths Cmax: 48-96hr 25mg/ml Method 2: ↑ decanoate dose over 4wks & ↓ po dose over 4-8wks as follows (accelerate taper for sx of EPS): t½: 6.8-9.6dy (single dose) ORAL DECANOATE (Administer Q 2 weeks) 15dy (14-100dy chronic administration) ORAL DOSE (mg/dy) ↓ DOSE OVER (wks) INITIAL DOSE (mg) TARGET DOSE (mg) DOSE OVER (wks) Steady State: 2mth (1.5-3mth) 5 4 6.25 6.25 0 Duration Action: 2wk (1-6wk) Elimination: Hepatic to inactive metabolites 10 4 6.25 12.5 4 Hemodialysis: Not dialyzable 20 8 6.25 12.5 4 30 8 6.25 25 4 40 8 6.25 25 4 Method 3: Admin equivalent decanoate dose Q2-3wks. -

Schizophrenia Care Guide
August 2015 CCHCS/DHCS Care Guide: Schizophrenia SUMMARY DECISION SUPPORT PATIENT EDUCATION/SELF MANAGEMENT GOALS ALERTS Minimize frequency and severity of psychotic episodes Suicidal ideation or gestures Encourage medication adherence Abnormal movements Manage medication side effects Delusions Monitor as clinically appropriate Neuroleptic Malignant Syndrome Danger to self or others DIAGNOSTIC CRITERIA/EVALUATION (PER DSM V) 1. Rule out delirium or other medical illnesses mimicking schizophrenia (see page 5), medications or drugs of abuse causing psychosis (see page 6), other mental illness causes of psychosis, e.g., Bipolar Mania or Depression, Major Depression, PTSD, borderline personality disorder (see page 4). Ideas in patients (even odd ideas) that we disagree with can be learned and are therefore not necessarily signs of schizophrenia. Schizophrenia is a world-wide phenomenon that can occur in cultures with widely differing ideas. 2. Diagnosis is made based on the following: (Criteria A and B must be met) A. Two of the following symptoms/signs must be present over much of at least one month (unless treated), with a significant impact on social or occupational functioning, over at least a 6-month period of time: Delusions, Hallucinations, Disorganized Speech, Negative symptoms (social withdrawal, poverty of thought, etc.), severely disorganized or catatonic behavior. B. At least one of the symptoms/signs should be Delusions, Hallucinations, or Disorganized Speech. TREATMENT OPTIONS MEDICATIONS Informed consent for psychotropic -

FDA Approved Drugs with Broad Anti-Coronaviral Activity Inhibit SARS-Cov-2 in Vitro
bioRxiv preprint doi: https://doi.org/10.1101/2020.03.25.008482; this version posted March 27, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. FDA approved drugs with broad anti-coronaviral activity inhibit SARS-CoV-2 in vitro Stuart Weston1, Rob Haupt1, James Logue1, Krystal Matthews1 and Matthew B. Frieman*1 1 - Department of Microbiology and Immunology, University of Maryland School of Medicine, 685 W. Baltimore St., Room 380, Baltimore, MD, 21201, USA *Corresponding author. Email: [email protected] Key words: SARS-CoV-2, nCoV-2019, COVID-19, drug repurposing, FDA approved drugs, antiviral therapeutics, pandemic, chloroquine, hydroxychloroquine AbstraCt SARS-CoV-2 emerged in China at the end of 2019 and has rapidly become a pandemic with over 400,000 recorded COVID-19 cases and greater than 19,000 recorded deaths by March 24th, 2020 (www.WHO.org) (1). There are no FDA approved antivirals or vaccines for any coronavirus, including SARS-CoV-2 (2). Current treatments for COVID-19 are limited to supportive therapies and off-label use of FDA approved drugs (3). Rapid development and human testing of potential antivirals is greatly needed. A potentially quicker way to test compounds with antiviral activity is through drug re-purposing (2, 4). Numerous drugs are already approved for use in humans and subsequently there is a good understanding of their safety profiles and potential side effects, making them easier to test in COVID-19 patients. -

NORPRAMIN® (Desipramine Hydrochloride Tablets USP)
NORPRAMIN® (desipramine hydrochloride tablets USP) Suicidality and Antidepressant Drugs Antidepressants increased the risk compared to placebo of suicidal thinking and behavior (suicidality) in children, adolescents, and young adults in short-term studies of major depressive disorder (MDD) and other psychiatric disorders. Anyone considering the use of NORPRAMIN or any other antidepressant in a child, adolescent, or young adult must balance this risk with the clinical need. Short-term studies did not show an increase in the risk of suicidality with antidepressants compared to placebo in adults beyond age 24; there was a reduction in risk with antidepressants compared to placebo in adults aged 65 and older. Depression and certain other psychiatric disorders are themselves associated with increases in the risk of suicide. Patients of all ages who are started on antidepressant therapy should be monitored appropriately and observed closely for clinical worsening, suicidality, or unusual changes in behavior. Families and caregivers should be advised of the need for close observation and communication with the prescriber. NORPRAMIN is not approved for use in pediatric patients. (See WARNINGS: Clinical Worsening and Suicide Risk, PRECAUTIONS: Information for Patients, and PRECAUTIONS: Pediatric Use.) DESCRIPTION NORPRAMIN® (desipramine hydrochloride USP) is an antidepressant drug of the tricyclic type, and is chemically: 5H-Dibenz[bƒ]azepine-5-propanamine,10,11-dihydro-N-methyl-, monohydrochloride. 1 Reference ID: 3536021 Inactive Ingredients The following inactive ingredients are contained in all dosage strengths: acacia, calcium carbonate, corn starch, D&C Red No. 30 and D&C Yellow No. 10 (except 10 mg and 150 mg), FD&C Blue No. 1 (except 25 mg, 75 mg, and 100 mg), hydrogenated soy oil, iron oxide, light mineral oil, magnesium stearate, mannitol, polyethylene glycol 8000, pregelatinized corn starch, sodium benzoate (except 150 mg), sucrose, talc, titanium dioxide, and other ingredients. -

PERPHENAZINE and AMITRIPTYLINE HYDROCHLORIDE- Perphenazine and Amitriptyline Hydrochloride Tablet, Film Coated Mylan Pharmaceuticals Inc
PERPHENAZINE AND AMITRIPTYLINE HYDROCHLORIDE- perphenazine and amitriptyline hydrochloride tablet, film coated Mylan Pharmaceuticals Inc. ---------- WARNING Increased Mortality in Elderly Patients with Dementia-Related Psychosis Elderly patients with dementia-related psychosis treated with antipsychotic drugs are at an increased risk of death. Analyses of seventeen placebo-controlled trials (modal duration of 10 weeks), largely in patients taking atypical antipsychotic drugs, revealed a risk of death in drug-treated patients of between 1.6 to 1.7 times the risk of death in placebo-treated patients. Over the course of a typical 10-week controlled trial, the rate of death in drug- treated patients was about 4.5%, compared to a rate of about 2.6% in the placebo group. Although the causes of death were varied, most of the deaths appeared to be either cardiovascular (e.g., heart failure, sudden death) or infectious (e.g., pneumonia) in nature. Observational studies suggest that, similar to atypical antipsychotic drugs, treatment with conventional antipsychotic drugs may increase mortality. The extent to which the findings of increased mortality in observational studies may be attributed to the antipsychotic drug as opposed to some characteristic(s) of the patients is not clear. Perphenazine and amitriptyline hydrochloride is not approved for the treatment of patients with dementia- related psychosis (see WARNINGS). Suicidality and Antidepressant Drugs Antidepressants increased the risk compared to placebo of suicidal thinking and behavior (suicidality) in children, adolescents and young adults in short-term studies of major depressive disorder (MDD) and other psychiatric disorders. Anyone considering the use of perphenazine and amitriptyline or any other antidepressant in a child, adolescent, or young adult must balance this risk with the clinical need. -

Detecting Depression and Anxiety in Older Adults
Detecting Depression and Anxiety in Older Adults Dr Nick Woodthorpe Depression in Older Adults Introduction to Depression • 1 in 5 older people living in the community • 2 in 5 living in care homes Causes of Depression • Painful events • Past depression or family history • Personality • Physical illness • Medicines • Drugs and Alcohol • Loneliness Particular Issues in Depression • Physical symptoms, for example, thyroid problems, heart disease or arthritis. • Long-term illness • Confusion and memory problems • A new sense of loneliness/lack of purpose Depression - Symptoms • 2 weeks • No hypomania or mania • No psychoactive drugs • No organic mental disorder Depression - Symptoms • Feeling low or sad (they may not report this) • Loss of interest in life • Low energy and tired for no reason Depression - Symptoms • Change in appetite and weight • Restlessness or psychomotor retardation • Anxious • Avoidance • Irritable • Sleep disturbance • Loss of confidence and self-esteem Depression - Symptoms • Hopelessness and worthlessness • Poor concentration • Panicky • Loss of libido • Sense of guilt • Suicidal thoughts • Delusions or hallucinations Depression - Diagnostic Categories • Mild • Moderate • Severe with or without psychosis • Recurrent Anxiety in Later Life Everyone gets anxious sometimes Everyone gets anxious sometimes Fight or Flight Response Fight or Flight Response Causes of anxiety in later life • Painful events • Stress • Past experiences- upbringing/Childhood • Personality • Physical illness • Medicines • Diet - caffeine, excess -

Association of Selective Serotonin Reuptake Inhibitors with the Risk for Spontaneous Intracranial Hemorrhage
Supplementary Online Content Renoux C, Vahey S, Dell’Aniello S, Boivin J-F. Association of selective serotonin reuptake inhibitors with the risk for spontaneous intracranial hemorrhage. JAMA Neurol. Published online December 5, 2016. doi:10.1001/jamaneurol.2016.4529 eMethods 1. List of Antidepressants for Cohort Entry eMethods 2. List of Antidepressants According to the Degree of Serotonin Reuptake Inhibition eMethods 3. Potential Confounding Variables Included in Multivariate Models eMethods 4. Sensitivity Analyses eFigure. Flowchart of Incident Antidepressant (AD) Cohort Definition and Case- Control Selection eTable 1. Crude and Adjusted Rate Ratios of Intracerebral Hemorrhage Associated With Current Use of SSRIs Relative to TCAs eTable 2. Crude and Adjusted Rate Ratios of Subarachnoid Hemorrhage Associated With Current Use of SSRIs Relative to TCAs eTable 3. Crude and Adjusted Rate Ratios of Intracranial Extracerebral Hemorrhage Associated With Current Use of SSRIs Relative to TCAs. eTable 4. Crude and Adjusted Rate Ratios of Intracerebral Hemorrhage Associated With Current Use of Antidepressants With Strong Degree of Inhibition of Serotonin Reuptake Relative to Weak eTable 5. Crude and Adjusted Rate Ratios of Subarachnoid Hemorrhage Associated With Current Use of Antidepressants With Strong Degree of Inhibition of Serotonin Reuptake Relative to Weak eTable 6. Crude and Adjusted Rate Ratios of Intracranial Extracerebral Hemorrhage Associated With Current Use of Antidepressants With Strong Degree of Inhibition of Serotonin Reuptake Relative to Weak This supplementary material has been provided by the authors to give readers additional information about their work. © 2016 American Medical Association. All rights reserved. Downloaded From: https://jamanetwork.com/ on 10/02/2021 eMethods 1. -

HALDOL Decanoate 50 (Haloperidol)
HALDOL® Decanoate 50 (haloperidol) HALDOL® Decanoate 100 (haloperidol) For IM Injection Only WARNING Increased Mortality in Elderly Patients with Dementia-Related Psychosis Elderly patients with dementia-related psychosis treated with antipsychotic drugs are at an increased risk of death. Analyses of seventeen placebo-controlled trials (modal duration of 10 weeks), largely in patients taking atypical antipsychotic drugs, revealed a risk of death in drug-treated patients of between 1.6 to 1.7 times the risk of death in placebo-treated patients. Over the course of a typical 10-week controlled trial, the rate of death in drug-treated patients was about 4.5%, compared to a rate of about 2.6% in the placebo group. Although the causes of death were varied, most of the deaths appeared to be either cardiovascular (e.g., heart failure, sudden death) or infectious (e.g., pneumonia) in nature. Observational studies suggest that, similar to atypical antipsychotic drugs, treatment with conventional antipsychotic drugs may increase mortality. The extent to which the findings of increased mortality in observational studies may be attributed to the antipsychotic drug as opposed to some characteristic(s) of the patients is not clear. HALDOL Decanoate is not approved for the treatment of patients with dementia-related psychosis (see WARNINGS). DESCRIPTION Haloperidol decanoate is the decanoate ester of the butyrophenone, HALDOL (haloperidol). It has a markedly extended duration of effect. It is available in sesame oil in sterile form for intramuscular (IM) injection. The structural formula of haloperidol decanoate, 4-(4-chlorophenyl)-1-[4-(4-fluorophenyl)-4-oxobutyl]-4 piperidinyl decanoate, is: Haloperidol decanoate is almost insoluble in water (0.01 mg/mL), but is soluble in most organic solvents. -

Schizophrenia Paper
The new england journal of medicine review article drug therapy Alastair J.J. Wood, M.D., Editor Schizophrenia Robert Freedman, M.D. From the Institute for Children’s Mental chizophrenia is a chronic, debilitating psychotic mental dis- Disorders, University of Colorado and the order that affects about 1 percent of people. A new generation of medications Veterans Affairs Medical Center, Denver. s Address reprint requests to Dr. Freedman and recent developments in neuropathology, brain imaging, and molecular ge- at the Department of Psychiatry, C-268-71, netics have led to a greater understanding of the pathophysiology of schizophrenia and University of Colorado Health Sciences to improved treatment. Nonetheless, it remains an enigmatic illness that places a sub- Center, Denver CO 80262, or at robert. [email protected]. stantial burden on patients, their families, and society. N Engl J Med 2003;349:1738-49. clinical characteristics Copyright © 2003 Massachusetts Medical Society. Schizophrenia has varied and ominous symptoms that generally begin in late adoles- cence or early adulthood and usually continue throughout life.1 Most patients have a history of behavioral dysfunction — primarily social and learning difficulties.2 Diagnos- tic features of schizophrenia include auditory hallucinations (generally voices that con- verse with or about the patient) and delusions (often the paranoid belief that external forces are conspiring against the patient). Patients may have some insight that the voices are internal thoughts and that the delusions cannot possibly be true, but these phenom- ena remain persistent and troubling. In addition to these overt psychotic, or “positive,” symptoms, various deficits, or “negative” symptoms, occur, including an inability to pay attention, the loss of a sense of pleasure, the loss of will or drive, disorganization or im- poverishment of thoughts and speech, flattening of affect, and social withdrawal (Ta- ble 1).