The Role of Hydrology in the Ecology of Cooper Creeþ Central Australia: Implications for the Flood Pulse Concept
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Painted Snippets the Newsletter of the Australian Painted Snipe Project
Painted Snippets The newsletter of the Australian Painted Snipe Project Volume 4 May 2012 A stunning female bathes herself, unperturbed by onlookers at Canberra’s Kelly Swamp. Photo: David Stowe None of us would have predicted the events Just add water which unfolded in the months that followed, It’s been an eventful 2 years since our last when one-in-10-year rains extended south edition of Painted Snippets hit the stands. After from the tropics and caused extensive flooding the 2008/09 summer, our concerns for the throughout 4 states, rejuvenating wetland and species were reinforced by a return of just 11 river systems throughout the Murray Darling, individuals observed across Australia. Bulloo-Bancannia and Lake Eyre Basins as well as across the Top End. Once the floods began to subside, observers Contents ventured out, discovering ephemeral wetlands which, in some cases, hadn’t been inundated in 1. Just Add Water 20 years! Soon enough, Australian Painted 2. Around the soggy grounds Snipe (APS) records started rolling in. With 5. Déjà vu 2005 conditions remaining wet throughout the year 6. A bird in the hand and Tropical Cyclone Yasi providing similar flows in early 2011, the last 2 years have seen 7. BirdLife Australia Wetland Birds Project over 400 individual APS1 recorded (Fig 2)across 8. iBis for your iPhone every state and territory except Tasmania, in 8. APS EPBC listing upgrade what has been true testament to the 9. Moolort Plains wetland project opportunistic nature of this enigmatic wader. 9. APS surveys. How to contribute to species conservation. -

Report to Office of Water Science, Department of Science, Information Technology and Innovation, Brisbane
Lake Eyre Basin Springs Assessment Project Hydrogeology, cultural history and biological values of springs in the Barcaldine, Springvale and Flinders River supergroups, Galilee Basin and Tertiary springs of western Queensland 2016 Department of Science, Information Technology and Innovation Prepared by R.J. Fensham, J.L. Silcock, B. Laffineur, H.J. MacDermott Queensland Herbarium Science Delivery Division Department of Science, Information Technology and Innovation PO Box 5078 Brisbane QLD 4001 © The Commonwealth of Australia 2016 The Queensland Government supports and encourages the dissemination and exchange of its information. The copyright in this publication is licensed under a Creative Commons Attribution 3.0 Australia (CC BY) licence Under this licence you are free, without having to seek permission from DSITI or the Commonwealth, to use this publication in accordance with the licence terms. You must keep intact the copyright notice and attribute the source of the publication. For more information on this licence visit http://creativecommons.org/licenses/by/3.0/au/deed.en Disclaimer This document has been prepared with all due diligence and care, based on the best available information at the time of publication. The department holds no responsibility for any errors or omissions within this document. Any decisions made by other parties based on this document are solely the responsibility of those parties. Information contained in this document is from a number of sources and, as such, does not necessarily represent government or departmental policy. If you need to access this document in a language other than English, please call the Translating and Interpreting Service (TIS National) on 131 450 and ask them to telephone Library Services on +61 7 3170 5725 Citation Fensham, R.J., Silcock, J.L., Laffineur, B., MacDermott, H.J. -

Ramsar Sites in Order of Addition to the Ramsar List of Wetlands of International Importance
Ramsar sites in order of addition to the Ramsar List of Wetlands of International Importance RS# Country Site Name Desig’n Date 1 Australia Cobourg Peninsula 8-May-74 2 Finland Aspskär 28-May-74 3 Finland Söderskär and Långören 28-May-74 4 Finland Björkör and Lågskär 28-May-74 5 Finland Signilskär 28-May-74 6 Finland Valassaaret and Björkögrunden 28-May-74 7 Finland Krunnit 28-May-74 8 Finland Ruskis 28-May-74 9 Finland Viikki 28-May-74 10 Finland Suomujärvi - Patvinsuo 28-May-74 11 Finland Martimoaapa - Lumiaapa 28-May-74 12 Finland Koitilaiskaira 28-May-74 13 Norway Åkersvika 9-Jul-74 14 Sweden Falsterbo - Foteviken 5-Dec-74 15 Sweden Klingavälsån - Krankesjön 5-Dec-74 16 Sweden Helgeån 5-Dec-74 17 Sweden Ottenby 5-Dec-74 18 Sweden Öland, eastern coastal areas 5-Dec-74 19 Sweden Getterön 5-Dec-74 20 Sweden Store Mosse and Kävsjön 5-Dec-74 21 Sweden Gotland, east coast 5-Dec-74 22 Sweden Hornborgasjön 5-Dec-74 23 Sweden Tåkern 5-Dec-74 24 Sweden Kvismaren 5-Dec-74 25 Sweden Hjälstaviken 5-Dec-74 26 Sweden Ånnsjön 5-Dec-74 27 Sweden Gammelstadsviken 5-Dec-74 28 Sweden Persöfjärden 5-Dec-74 29 Sweden Tärnasjön 5-Dec-74 30 Sweden Tjålmejaure - Laisdalen 5-Dec-74 31 Sweden Laidaure 5-Dec-74 32 Sweden Sjaunja 5-Dec-74 33 Sweden Tavvavuoma 5-Dec-74 34 South Africa De Hoop Vlei 12-Mar-75 35 South Africa Barberspan 12-Mar-75 36 Iran, I. R. -

DUCK HUNTING in VICTORIA 2020 Background
DUCK HUNTING IN VICTORIA 2020 Background The Wildlife (Game) Regulations 2012 provide for an annual duck season running from 3rd Saturday in March until the 2nd Monday in June in each year (80 days in 2020) and a 10 bird bag limit. Section 86 of the Wildlife Act 1975 enables the responsible Ministers to vary these arrangements. The Game Management Authority (GMA) is an independent statutory authority responsible for the regulation of game hunting in Victoria. Part of their statutory function is to make recommendations to the relevant Ministers (Agriculture and Environment) in relation to open and closed seasons, bag limits and declaring public and private land open or closed for hunting. A number of factors are reviewed each year to ensure duck hunting remains sustainable, including current and predicted environmental conditions such as habitat extent and duck population distribution, abundance and breeding. This review however, overlooks several reports and assessments which are intended for use in managing game and hunting which would offer a more complete picture of habitat, population, abundance and breeding, we will attempt to summarise some of these in this submission, these include: • 2019-20 Annual Waterfowl Quota Report to the Game Licensing Unit, New South Wales Department of Primary Industries • Assessment of Waterfowl Abundance and Wetland Condition in South- Eastern Australia, South Australian Department for Environment and Water • Victorian Summer waterbird Count, 2019, Arthur Rylah Institute for Environmental Research As a key stakeholder representing 17,8011 members, Field & Game Australia Inc. (FGA) has been invited by GMA to participate in the Stakeholder Meeting and provide information to assist GMA brief the relevant Ministers, FGA thanks GMA for this opportunity. -

Management Plan for the South Australian Lake Eyre Basin Fisheries
MANAGEMENT PLAN FOR THE SOUTH AUSTRALIAN LAKE EYRE BASIN FISHERIES Part 1 – Commercial and recreational fisheries Part 2 – Yandruwandha Yawarrawarrka Aboriginal traditional fishery Approved by the Minister for Agriculture, Food and Fisheries pursuant to section 44 of the Fisheries Management Act 2007. Hon Gail Gago MLC Minister for Agriculture, Food and Fisheries 1 March 2013 Page 1 of 118 PIRSA Fisheries & Aquaculture (A Division of Primary Industries and Regions South Australia) GPO Box 1625 ADELAIDE SA 5001 www.pir.sa.gov.au/fisheries Tel: (08) 8226 0900 Fax: (08) 8226 0434 © Primary Industries and Regions South Australia 2013 Disclaimer: This management plan has been prepared pursuant to the Fisheries Management Act 2007 (South Australia) for the purpose of the administration of that Act. The Department of Primary Industries and Regions SA (and the Government of South Australia) make no representation, express or implied, as to the accuracy or completeness of the information contained in this management plan or as to the suitability of that information for any particular purpose. Use of or reliance upon information contained in this management plan is at the sole risk of the user in all things and the Department of Primary Industries and Regions SA (and the Government of South Australia) disclaim any responsibility for that use or reliance and any liability to the user. Copyright Notice: This work is copyright. Copyright in this work is owned by the Government of South Australia. Apart from any use permitted under the Copyright Act 1968 (Commonwealth), no part of this work may be reproduced by any process without written permission of the Government of South Australia. -

Heritage of the Birdsville and Strzelecki Tracks
Department for Environment and Heritage Heritage of the Birdsville and Strzelecki Tracks Part of the Far North & Far West Region (Region 13) Historical Research Pty Ltd Adelaide in association with Austral Archaeology Pty Ltd Lyn Leader-Elliott Iris Iwanicki December 2002 Frontispiece Woolshed, Cordillo Downs Station (SHP:009) The Birdsville & Strzelecki Tracks Heritage Survey was financed by the South Australian Government (through the State Heritage Fund) and the Commonwealth of Australia (through the Australian Heritage Commission). It was carried out by heritage consultants Historical Research Pty Ltd, in association with Austral Archaeology Pty Ltd, Lyn Leader-Elliott and Iris Iwanicki between April 2001 and December 2002. The views expressed in this publication are not necessarily those of the South Australian Government or the Commonwealth of Australia and they do not accept responsibility for any advice or information in relation to this material. All recommendations are the opinions of the heritage consultants Historical Research Pty Ltd (or their subconsultants) and may not necessarily be acted upon by the State Heritage Authority or the Australian Heritage Commission. Information presented in this document may be copied for non-commercial purposes including for personal or educational uses. Reproduction for purposes other than those given above requires written permission from the South Australian Government or the Commonwealth of Australia. Requests and enquiries should be addressed to either the Manager, Heritage Branch, Department for Environment and Heritage, GPO Box 1047, Adelaide, SA, 5001, or email [email protected], or the Manager, Copyright Services, Info Access, GPO Box 1920, Canberra, ACT, 2601, or email [email protected]. -
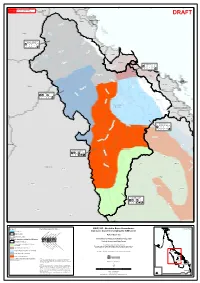
GWQ1203 - Burdekin Basin Groundwater Location Map Watercourse Cainozoic Deposits Overlying the GAB Zones
144°0'0"E 145°0'0"E 146°0'0"E 147°0'0"E 148°0'0"E 149°0'0"E Tully 18°0'0"S This map must not be used for marine navigation. 18°0'0"S Comprehensive and updated navigation WARNING information should be obtained from published Mitchell hydrographic charts. Murray DRAFT Hinchinbrook Island Herbert r ve Ri ekin urd B Ingham ! er iv D R ry Gilbert ry Ri D v er r e iv 19°0'0"S Scattered Remnants Northern R 19°0'0"S g Burdekin Headwaters in n n R u Black Townsville ! er r iv e R v rke i la R C r ta S Ross Broken River Ayr ! er ver Riv i ry lt R Haughton go Basa e r Don and Bogie Coastal Area G h t r u ve o i ton R S h ug Ha Bowen ! 20°0'0"S 20°0'0"S r Charters Towers e v ! i Don R B n o o g i D e Riv er Bur dek in R ive r Whitsunday Island Proserpine B owe Proserpine Flinders n ! er R Riv iver e sp a R amp C o Don River l l s to n R iv C e a r North West Suttor Catchment pe R r iv ve e i r R r o t t u S r e Broken v R i iv S e R e r llh L e e i O'Connell p im t t a l R e C iv er B o w e B u r d ekin n B u r d ekin R 21°0'0"S i 21°0'0"S v e BB a a s s in in r B ro Mackay ke ! n River Pioneer Plane Sarina ! Eastern Weathered Su ttor R Cainozoic Remnants Belyandor River iv e er iv R o d n lya e B r e v i R o d Moranbah n ! a ly 22°0'0"S e 22°0'0"S l R B ae iver ich Carm r ive R Diamantina s r o n n o C Styx Belyando River Saline Tertiary Sediments Dysart ! Is a a c R iv B Clermont Middlemount er e ! ! l y an do Riv e r er iv Cooper Creek R ie nz Aramac e ! k c a M 23°0'0"S 23°0'0"S Capella -

Studies of Ephydrinae (Diptera: Ephydridae), IV: Revision of the Australian Species of Subgenus Neoscatella Malloch
Studies of Ephydrinae (Diptera: Ephydridae), IV: Revision of the Australian Species of Subgenus Neoscatella Malloch WAYNE N. MATHIS and WILLIS W. WIRTH SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 325 SERIAL PUBLICATIONS OF THE SMITHSONIAN INSTITUTION The emphasis upon publications as a means of diffusing knowledge was expressed by the first Secretary of the Smithsonian Institution. In his formal plan for the Insti- tution, Joseph Henry articulated a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This keynote of basic research has been adhered to over the years in the issuance of thousands of titles in serial publications under the Smithsonian imprint, com- mencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Annals of Flight Smithsonian Contributions to Anthropology Smithsonian Contributions to Astrophysics Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoology Smithsonian Studies in History and Technology In these series, the Institution publishes original articles and monographs dealing with the research and collections of its several museums and offices and of professional colleagues at other institutions of learning. These papers report newly acquired facts, synoptic interpretations of data, or original theory in specialized fields. These pub- lications are distributed by mailing lists to libraries, laboratories, and other interested institutions and specialists throughout the world. Individual copies may be obtained from the Smithsonian Institution Press as long as stocks are available. -

Innamincka Regional Reserve Draft Management Plan 2017
Innamincka Regional Reserve Draft Management Plan 2017 Recognising the cultural and interconnected nature of Innamincka Regional Reserve, and working together towards sustainable land use Your views are important A management plan for the Innamincka Regional Reserve is being prepared to ensure the long term protection of the regional reserve’s natural values and advance spiritual, cultural, social and economic opportunities for the traditional custodians – the Yandruwandha people and the Yawarrawarrka people. The Innamincka Regional Reserve Draft Management Plan is now released for public consultation. Members of the community are encouraged to express their views on the future management of this regional reserve. A final plan will be prepared in response to submissions received on this draft plan. Once prepared, the final plan will be forwarded to the Minister for Sustainability, Environment and Conservation for consideration, together with a detailed analysis of submissions received. Notice of the adoption of the final plan will be published in the Government Gazette and the final Innamincka Regional Reserve Management Plan will be made available at: www.environment.sa.gov.au/parkmanagement. I encourage you to make a submission on this draft plan. John Schutz Director of National Parks and Wildlife Cultural Sensitivity Warning Aboriginal people are warned that this publication may contain culturally sensitive material. 1 Developing this draft plan This draft management plan was developed by the Department of Environment, Water and Natural Resources (DEWNR) with advice from the Yandruwandha Yawarrawarrka Parks Advisory Committee. It draws on feedback received in response to a stakeholder workshop and a discussion paper that was released to the public in 2015. -

Innamincka/Cooper Creek State Heritage Area Innamincka/Cooper Creek Was Declared a State Heritage Area on 16 May 1985
Innamincka/Cooper Creek State Heritage Area Innamincka/Cooper Creek was declared a State Heritage Area on 16 May 1985. HISTORY Innamincka and Cooper Creek have a rich history. The area has also been associated with many other facets of the state's history since colonial settlement. Names such as Charles Sturt, Sidney Kidman, Cordillo Downs, the Australian Inland Mission and the Reverend John Flynn are intrinsically woven into the stories of Innamincka and the Cooper Creek. Its significance for Aboriginal people spans thousands of years as a trade route and a source of abundant food and fresh water. The area's Aboriginal history also includes significant contacts with explorers, pastoralists and settlers. Many sources believe that the name 'Innamincka' comes from two Aboriginal words meaning ' your shelter' or 'your home'. European contact with the region came first with the explorers and later with the establishment of the pastoral industry, transport routes and service centres. Cooper Creek was named by Captain Charles Sturt on 13 October 1845, when he crossed the watercourse at a point approximately 24 kilometres west of the current Innamincka township. The water was very low at the time, hence his use of the term 'creek' rather than 'river' to describe what often becomes a deep torrent of water. Charles Cooper, after whom the waterway was named, was a South Australian judge - later Chief Justice Sir Charles Cooper. On the same expedition, Sturt also named the Strzelecki Desert after the eccentric Polish explorer, Paul Edmund de Strzelecki, who was the first European to climb and name Mount Kosciusko. -

100 the SOUTH-WEST CORNER of QUEENSLAND. (By S
100 THE SOUTH-WEST CORNER OF QUEENSLAND. (By S. E. PEARSON). (Read at a meeting of the Historical Society of Queensland, August 27, 1937). On a clear day, looking westward across the channels of the Mulligan River from the gravelly tableland behind Annandale Homestead, in south western Queensland, one may discern a long low line of drift-top sandhills. Round more than half the skyline the rim of earth may be likened to the ocean. There is no break in any part of the horizon; not a landmark, not a tree. Should anyone chance to stand on those gravelly rises when the sun was peeping above the eastem skyline they would witness a scene that would carry the mind at once to the far-flung horizons of the Sahara. In the sunrise that western region is overhung by rose-tinted haze, and in the valleys lie the purple shadows that are peculiar to the waste places of the earth. Those naked, drift- top sanddunes beyond the Mulligan mark the limit of human occupation. Washed crimson by the rising sun they are set Kke gleaming fangs in the desert's jaws. The Explorers. The first white men to penetrate that line of sand- dunes, in south-western Queensland, were Captain Charles Sturt and his party, in September, 1845. They had crossed the stony country that lies between the Cooper and the Diamantina—afterwards known as Sturt's Stony Desert; and afterwards, by the way, occupied in 1880, as fair cattle-grazing country, by the Broad brothers of Sydney (Andrew and James) under the run name of Goyder's Lagoon—and the ex plorers actually crossed the latter watercourse with out knowing it to be a river, for in that vicinity Sturt describes it as "a great earthy plain." For forty miles one meets with black, sundried soil and dismal wilted polygonum bushes in a dry season, and forty miles of hock-deep mud, water, and flowering swamp-plants in a wet one. -

FOIR-Coongie-Road-Survey-Project
COONGIE ROAD BIRDS, MAMMALS & VEGETATION SURVEY 2014 A project undertaken by the Friends of the Innamincka Reserves Dune near Coongie Road, Innamincka Regional Reserve i REPORT ON THE COONGIE ROAD BIRDS, MAMMALS & VEGETATION SURVEY 2014 CONTENTS Page INTRODUCTION 1 A. Project coordinator and field team 1 B. Background 1 C. Approach 2 D. Objectives 2 E. Programme of research 2 METHODS 3 RESULTS AND DISCUSSION 6 A. Bird survey data 6 B. Habitats 9 C. Flora 11 D. Mammals 13 E. Reptiles and amphibians 15 F. Threats and potential impacting factors 15 G. Archeological sites 18 CONCLUSIONS 19 APPENDIX I - Location of Census Stops 20 APPENDIX II - Transect Bird Data 23 APPENDIX III - Photographic and Habitat Records 27 APPENDIX IV - Using a GPS to Navigate a Transect 49 ii REPORT ON THE COONGIE ROAD BIRDS, MAMMALS & VEGETATION SURVEY 2014 INTRODUCTION A. PROJECT COORDINATOR AND FIELD TEAM Coordinator: Kate Buckley Team Leaders: Euan Moore, Jenny Rolland, Rose Treilibs, Vern Treilibs Field Team: Daphne Hards, Sonja Ross, Karen and Geoff Russell, Jen and Len Kenna, Barbara and Peter Bansemer, Fae and Jim Trueman, Jan and Ray Hutchinson In 2014 this project was carried out as a volunteer activity by members of the Friends of Innamincka Reserves (FOIR). There was no external funding for the project. B. BACKGROUND The Coongie Road extends from Innamincka north- west to Malkumba-Coongie Lakes NP via Kudriemitchie. It passes through a range of habitat types from dry grasslands to wetlands. While average rainfall is low (177 mm per annum), the Innamincka area is in a region of maximum rainfall variability for Australia.