Complete Issue
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

New Oriental Education & Technology Group Inc
Table of Contents UNITED STATES SECURITIES AND EXCHANGE COMMISSION Washington, D.C. 20549 FORM 20-F (Mark One) ☐ REGISTRATION STATEMENT PURSUANT TO SECTION 12(B) OR 12(G) OF THE SECURITIES EXCHANGE ACT OF 1934 OR ☒ ANNUAL REPORT PURSUANT TO SECTION 13 OR 15(D) OF THE SECURITIES EXCHANGE ACT OF 1934 For the fiscal year ended May 31, 2011. OR ☐ TRANSITION REPORT PURSUANT TO SECTION 13 OR 15(D) OF THE SECURITIES EXCHANGE ACT OF 1934 OR ☐ SHELL COMPANY REPORT PURSUANT TO SECTION 13 OR 15(D) OF THE SECURITIES EXCHANGE ACT OF 1934 Date of event requiring this shell company report For the transition period from to Commission file number: 001-32993 NEW ORIENTAL EDUCATION & TECHNOLOGY GROUP INC. (Exact name of Registrant as specified in its charter) N/A (Translation of Registrant’s name into English) Cayman Islands (Jurisdiction of incorporation or organization) No. 6 Hai Dian Zhong Street Haidian District, Beijing 100080 The People’s Republic of China (Address of principal executive offices) Louis T. Hsieh, President and Chief Financial Officer Tel: +(86 10) 6260-5566 E-mail: [email protected] Fax: +(86 10) 6260-5511 No. 6 Hai Dian Zhong Street Haidian District, Beijing 100080 The People’s Republic of China (Name, Telephone, E-mail and/or Facsimile number and Address of Company Contact Person) Securities registered or to be registered pursuant to Section 12(b) of the Act: Title of Each Class Name of Exchange on Which Registered American depositary shares, each representing one New York Stock Exchange common share* Common shares, par value US$0.01 per share New York Stock Exchange** * Effective August 18, 2011, the ratio of ADSs to our common shares was changed from one ADS representing four common shares to one ADS representing one common share. -

Table of Codes for Each Court of Each Level
Table of Codes for Each Court of Each Level Corresponding Type Chinese Court Region Court Name Administrative Name Code Code Area Supreme People’s Court 最高人民法院 最高法 Higher People's Court of 北京市高级人民 Beijing 京 110000 1 Beijing Municipality 法院 Municipality No. 1 Intermediate People's 北京市第一中级 京 01 2 Court of Beijing Municipality 人民法院 Shijingshan Shijingshan District People’s 北京市石景山区 京 0107 110107 District of Beijing 1 Court of Beijing Municipality 人民法院 Municipality Haidian District of Haidian District People’s 北京市海淀区人 京 0108 110108 Beijing 1 Court of Beijing Municipality 民法院 Municipality Mentougou Mentougou District People’s 北京市门头沟区 京 0109 110109 District of Beijing 1 Court of Beijing Municipality 人民法院 Municipality Changping Changping District People’s 北京市昌平区人 京 0114 110114 District of Beijing 1 Court of Beijing Municipality 民法院 Municipality Yanqing County People’s 延庆县人民法院 京 0229 110229 Yanqing County 1 Court No. 2 Intermediate People's 北京市第二中级 京 02 2 Court of Beijing Municipality 人民法院 Dongcheng Dongcheng District People’s 北京市东城区人 京 0101 110101 District of Beijing 1 Court of Beijing Municipality 民法院 Municipality Xicheng District Xicheng District People’s 北京市西城区人 京 0102 110102 of Beijing 1 Court of Beijing Municipality 民法院 Municipality Fengtai District of Fengtai District People’s 北京市丰台区人 京 0106 110106 Beijing 1 Court of Beijing Municipality 民法院 Municipality 1 Fangshan District Fangshan District People’s 北京市房山区人 京 0111 110111 of Beijing 1 Court of Beijing Municipality 民法院 Municipality Daxing District of Daxing District People’s 北京市大兴区人 京 0115 -

New Oriental Education & Technology Group Inc
Table of Contents UNITED STATES SECURITIES AND EXCHANGE COMMISSION Washington, D.C. 20549 FORM 20-F (Mark One) ☐ REGISTRATION STATEMENT PURSUANT TO SECTION 12(B) OR 12(G) OF THE SECURITIES EXCHANGE ACT OF 1934 OR ☒ ANNUAL REPORT PURSUANT TO SECTION 13 OR 15(D) OF THE SECURITIES EXCHANGE ACT OF 1934 For the fiscal year ended May 31, 2012. OR ☐ TRANSITION REPORT PURSUANT TO SECTION 13 OR 15(D) OF THE SECURITIES EXCHANGE ACT OF 1934 OR ☐ SHELL COMPANY REPORT PURSUANT TO SECTION 13 OR 15(D) OF THE SECURITIES EXCHANGE ACT OF 1934 Date of event requiring this shell company report For the transition period from to Commission file number: 001-32993 NEW ORIENTAL EDUCATION & TECHNOLOGY GROUP INC. (Exact name of Registrant as specified in its charter) N/A (Translation of Registrant’s name into English) Cayman Islands (Jurisdiction of incorporation or organization) No. 6 Hai Dian Zhong Street Haidian District, Beijing 100080 People’s Republic of China (Address of principal executive offices) Louis T. Hsieh, President and Chief Financial Officer Tel: +(86 10) 6260-5566 E-mail: [email protected] Fax: +(86 10) 6260-5511 No. 6 Hai Dian Zhong Street Haidian District, Beijing 100080 People’s Republic of China (Name, Telephone, E-mail and/or Facsimile number and Address of Company Contact Person) Securities registered or to be registered pursuant to Section 12(b) of the Act: Title of Each Class Name of Exchange on Which Registered American depositary shares, each representing one common share* New York Stock Exchange Common shares, par value US$0.01 per share** New York Stock Exchange * Effective August 18, 2011, the ratio of ADSs to our common shares was changed from one ADS representing four common shares to one ADS representing one common share. -
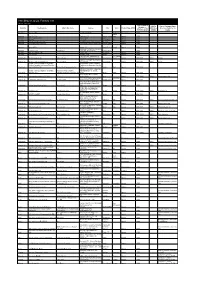
Levi Strauss & Co. Factory List
Levi Strauss & Co. Factory List Published : August 2020 Total Number of LS&Co. Parent Company Name Employees Country Factory name Alternative Name Address City State Product Type (TOE) Initiatives (Licensee factories are (Workers, Staff, (WWB) blank) Contract Staff) Argentina Accecuer SA Juan Zanella 4656 Caseros Dong Nai Accessories <1000 Tanmil Argentina Best Sox S.A. Charlone 1446 Capital Federal Nadu Apparel <1000 Argentina Estex Argentina S.R.L. Superi, 3530 Caba Apparel <1000 Argentina Gitti SRL Italia 4043 Mar del Plata TN Apparel <1000 Argentina Manufactura Arrecifes S.A. Ruta Nacional 8, Kilometro 178 Arrecifes Apparel <1000 Argentina Procesadora Serviconf SRL Gobernardor Ramon Castro 4765 Vicente Lopez Apparel <1000 Argentina Spring S.R.L. Darwin, 173 Capital Federal Punjab Apparel <1000 Asamblea (101) #536, Villa Lynch Argentina TEXINTER S.A. Texinter S.A. B1672AIB, Buenos Aires Buenos Aires <1000 Argentina Underwear M&S, S.R.L Levalle 449 Avellaneda Apparel <1000 Argentina Vira Offis S.A. Virasoro, 3570 Rosario Apparel <1000 Plot # 246-249, Adamjee EPZ, Rio Grande Bangladesh Ananta Apparels Ltd. Nazmul Hoque Shiddirgonj, Narayangonj-1431 Narayangonj do Sul Apparel 1000-5000 WWB Ananta Nayabari, Kanchpur, Sonargaon, Bangladesh Ananta Denim Technology Ltd. Md. Ziaul Alam Narayangonj. Narayanganj Apparel 1000-5000 WWB Ananta Ayesha Clothing Company Ltd (Ayesha Bangobandhu Road, Tongabari, Clothing Company Ltd,Hamza Trims Ltd, Gazirchat Alia Madrasha, Ashulia, Bangladesh Hamza Clothing Ltd) Ayesha Clothing Company Ltd( Dhaka Dhaka Sindh Apparel 1000-5000 Jamgora, Post Office : Gazirchat Ayesha Clothing Company Ltd (Ayesha Ayesha Clothing Company Alia Madrasha, P.S : Savar, Bangladesh Washing Ltd.) Ltd(Unit-1)d (Ayesha Washing Ltd) Dhaka Dhaka Sindh Apparel 1000-5000 Khejur Bagan, Bara Ashulia, Bangladesh Cosmopolitan Industries PVT Ltd CIPL Savar, Dhaka Savar Apparel 5000-10000 WWB Epic Designers Ltd 1612, South Salna, Salna Bazar, TAMILNAD Bangladesh Cutting Edge Industries Ltd. -

SUNAC CHINA HOLDINGS LIMITED 融創中國控股有限公司 (Incorporated in the Cayman Islands with Limited Liability) (Stock Code: 01918)
Hong Kong Exchanges and Clearing Limited and The Stock Exchange of Hong Kong Limited take no 14.88 responsibility for the contents of this announcement, make no representation as to its accuracy or completeness and expressly disclaim any liability whatsoever for any loss howsoever arising from or in reliance upon the whole or any part of the contents of this announcement. SUNAC CHINA HOLDINGS LIMITED 融創中國控股有限公司 (Incorporated in the Cayman Islands with limited liability) (Stock Code: 01918) (1) VERY SUBSTANTIAL ACQUISITION ENTERING INTO OF FRAMEWORK AGREEMENT IN RELATION TO THE COOPERATION OF TARGET PROJECT COMPANIES AND TARGET HOTEL ASSETS AND (2) RESUMPTION OF TRADING IN SHARES THE COOPERATION The Board is pleased to announce that, on 10 July 2017, Sunac Real Estate, an indirect wholly-owned subsidiary of the Company as the buyer, and Dalian Wanda Commercial Properties as the seller entered into the Framework Agreement, pursuant to which the Buyer agreed to acquire, and the Seller agreed to dispose of 91% equity interest of the 13 cultural and tourism project companies in the PRC (i.e. the Target Project Companies) and 100% interest of the 76 city hotels (i.e. the Target Hotel Assets) at the consideration of approximately RMB29,575,000,000 and RMB33,595,260,800 respectively. Thus, the total consideration for the Cooperation is approximately RMB63,170,260,800. Before completion of the Cooperation, the Seller held 100% equity interest of the Target Project Companies and 100% interest of the Target Hotel Assets. After completion of the Cooperation, the Buyer will hold 91% equity interest of the Target Project Companies and 100% interest of the Target Hotel Assets, while the Seller will continue to hold 9% equity interest of the Target Project Companies. -

Yichang to Badong Expressway Project
IPP307 v2 World Bank Financed Highway Project Yiba Expressway in Hubei·P. R. China YBE_05 Public Disclosure Authorized Public Disclosure Authorized Yichang to Badong Expressway Project Social Assessment Report Final Public Disclosure Authorized Public Disclosure Authorized The World Bank Financed Project Office of HPCD Social Survey Research Center of Peking University 22 September 2008 1 Preface Entrusted by the World Bank Financed Project Execution Office (PEO) under the Hubei Provincial Communications Department (HPCD), the Social Survey Research Center of Peking University (SSRCPKU) conducted an independent assessment on the “Project of the Stretch from Yichang to Badong of the Highway from Shanghai to Chengdu”. The Yiba stretch of the highway from Shanghai to Chengdu is lying in the west of Hubei Province which is at the joint of middle reaches and upper reaches of the Yangtze River. The project area administratively belongs to Yiling District Yichang City, Zigui County, Xingshang County and Badong County of Shien Tujia & Miao Autonomous Prefecture. It adjoins Jianghan Plain in the east, Chongqing City in the west, Yangtze River in the south and Shengnongjia Forest, Xiangfan City etc in the north. The highway, extending 173 km, begins in Baihe, connecting Jingyi highway, and ends up in Badong County in the joint of Hubei and Sichuan, joining Wufeng highway in Chongqing. Under the precondition of sticking to the World Bank’s policy, the social assessment is going to make a judgment of the social impact exerted by the project, advance certain measures, and in the meanwhile bring forward supervision and appraisement system. During July 1st and 9th, 2007, the assessment team conducted the social investigation in Yiling District Yichang City, Zigui County, XingshanCounty and Badong County. -

World Bank Financed Hubei Yichang Three Gorges Modern Logistics Center Infrastructure Project
SFG2362 G.H.P.Z.J.Z. No. 2606 Public Disclosure Authorized World Bank Financed Hubei Yichang Three Gorges Modern Logistics Center Infrastructure Project Public Disclosure Authorized Environmental and Social Impact Assessment Report (For Appraisal) Public Disclosure Authorized Public Disclosure Authorized Central-Southern Safety & Environment Technology Institute Co. Ltd. July, 2016 World Bank Financed Hubei Yichang Three Gorges Modern Name of the Project Logistics Center Infrastructure Project Central-Southern Safety & Environment Technology Institute Assessed by Co. Ltd. Legal representative QIAN Bin Task Team leader LIU Zhe Vice-Task Team Leader SHE Jian Team Member KOU Xueyong, LUO Feng, ZHANG Qi Reviewed and Approved by: ZHANG Bin Table of Contents 1 INTRODUCTION ......................................................................................................................... - 1 - 1.1 BACKGROUND ................................................................................................................................ - 1 - 1.2 REGULATORY AND LEGAL FRAMEWORK .......................................................................................... - 1 - 1.2.1 Laws and regulations .................................................................................................................- 1 - 1.2.2 Administration regulations ........................................................................................................- 3 - 1.2.3 Technical specifications ..............................................................................................................- -

Major Transaction in Relation to the Acquisition of Hotel Assets
Hong Kong Exchanges and Clearing Limited and The Stock Exchange of Hong Kong Limited take no responsibility for the contents of this announcement, make no representation as to its accuracy or completeness and expressly disclaim any liability whatsoever for any loss howsoever arising from or in reliance upon the whole or any part of the contents of this announcement. (a joint stock limited company incorporated in the People’s Republic of China with limited liability) (Stock Code: 2777) MAJOR TRANSACTION IN RELATION TO THE ACQUISITION OF HOTEL ASSETS THE ACQUISITION On 19 July 2017 (after trading hours), the Company as the buyer, and Dalian Wanda as the seller entered into the Agreement, pursuant to which the Company conditionally agreed to acquire, and Dalian Wanda conditionally agreed to dispose of, the interest in 76 city hotels and a 70% interest in Yantai Wanda at the consideration of approximately RMB19,906,390,000. IMPLICATIONS UNDER THE LISTING RULES As one or more applicable percentage ratios under the Rule 14.07 of the Listing Rules in respect of the Acquisition exceed 25% and all of the applicable percentage ratios are less than 100%, the Acquisition constitutes a major transaction of the Company under Chapter 14 of the Listing Rules and is therefore subject to the reporting, announcement, circular and shareholders’ approval requirements under Chapter 14 of the Listing Rules. An extraordinary general meeting will be convened and held for the Shareholders to approve the Agreement and the transactions contemplated thereunder. To the best knowledge, information and belief of the Directors, after making all reasonable enquiries, no Shareholder or any of its associate has any material interest in the Agreement and the transaction contemplated thereunder and is required to abstain from voting. -

Improved Grey Clustering Method in Risk Zonation of Mountain Flash Flood
International Journal of Environmental & Agriculture Research (IJOEAR) ISSN:[2454-1850] [Vol-4, Issue-5, May- 2018] Improved grey clustering method in risk zonation of mountain flash flood disaster Qiong Li1, Jianzhong Zhou2*, Jiaming Cai3, Huaiwei Sun4 1School of Mathematics and Physics, Hubei Polytechnic University, Huangshi 435003, China 1,2,3,4College of Hydropower and Information Engineering, Huazhong University of Science and Technology, Wuhan 430074, China Abstract— Flash floods are considered one of the worst weather-related natural disasters. Flash floods are dangerous because they are sudden and highly unpredictable. Identification of the locations of high-risk areas has a major effect on the improvement of flash flood disaster control and prevention. Earlier work conducted on flood disaster risk zonation was commonly based on Digital Elevation Mode (DEM) data and statistical yearbook data and used an index, such as rainfall, topography, slope, or river distribution, with the analytic hierarchy process (AHP) method to determine the weighting. In this method, the final regional risk map was created by using ArcGIS map algebra superposition. In the present study, an improved gray clustering method is put forward to improve the comprehensive evaluation of the risk of mountain flash flood disasters by constructing the exponential whitening function and by using the information entropy weight method, which produces results that are more accurate and more reliable than those of the traditional method. This improved method can make full use of the limited information available, improving not only the resolution but also the influence of the subjective method, and produces more objective and accurate evaluation results. We obtain the risk degree by combining the information entropy weight and improved whitening function approaches in a gray clustering methodology. -

Annual Report
Important Notice: 1. The Board, the Supervisory Committee and the Directors, members of the Supervisory Committee and senior management of the Company warrant that in respect of the information contained in 2020 Annual Report (the “Report”, or “Annual Report”), there are no misrepresentations, misleading statements or material omission, and individually and collectively accept full responsibility for the authenticity, accuracy and completeness of the information contained in the Report. 2. The Report has been approved by the sixth meeting of the 19th session of the Board (the “Meeting”) convened on 30 March 2021. Mr. XIN Jie and Mr. TANG Shaojie, both being Non-executive Directors, did not attend the Meeting due to business engagement, and had authorised Mr. LI Qiangqiang, also a Non-executive Director, to attend the Meeting and executed voting rights on their behalf. All other Directors attended the Meeting in person. 3. The Company’s proposal on dividend distribution for the year of 2020: Based on the total share capital on the equity registration date when dividends are paid, the total amount of cash dividends proposed for distribution for 2020 will be RMB14,522,165,251.25 (inclusive of tax), accounting for 34.98% of the net profit attributable to equity shareholders of the Company for 2020, without any bonus shares or transfer of equity reserve to the share capital. Based on the Company’s total number of 11,617,732,201 shares at the end of 2020, a cash dividend of RMB12.5 (inclusive of tax) will be distributed for each 10 shares. If any circumstances, such as issuance of new shares, share repurchase or conversion of any convertible bonds into share capital before the record date for dividend distribution, results in the changes in our total number of shares on record date for dividend distribution, dividend per share shall be adjusted accordingly on the premise that the total dividends amount remains unchanged. -

A Spatial Distribution Equilibrium Evaluation of Health Service Resources at Community Grid Scale in Yichang, China
sustainability Article A Spatial Distribution Equilibrium Evaluation of Health Service Resources at Community Grid Scale in Yichang, China Jingyuan Chen 1, Yuqi Bai 1,2,3,* , Pei Zhang 4,*, Jingyuan Qiu 2, Yichun Hu 5, Tianhao Wang 6, Chengzhong Xu 4 and Peng Gong 2,3 1 Faculty of Innovation and Design, City University of Macau, Taipa, Macau 999078, China; [email protected] 2 Department of Earth System Science, Tsinghua University, Beijing 100084, China; [email protected] (J.Q.); [email protected] (P.G.) 3 Center for Healthy Cities, Institute for China Sustainable Urbanization, Tsinghua University, Beijing 100875, China 4 Center for Disease Control and Prevention, Yichang 443000, China; [email protected] 5 Smart City Construction Office, Yichang 443000, China; [email protected] 6 Big Data Management Center, Yichang 443000, China; [email protected] * Correspondence: [email protected] (Y.B.); [email protected] (P.Z.) Received: 11 November 2019; Accepted: 16 December 2019; Published: 19 December 2019 Abstract: Whether the supplies of health services and related facilities meet the demand is a critical issue when developing healthy cities. The importance of health services and related facilities in public health promotion has been adequately proved. However, since the community population and resource data are usually available at the scale of an administrative region; it is very difficult to perform further fine-scaled spatial distribution equilibrium evaluation studies. Such kinds of activities are highly expected for precise urban planning and management. Yichang is located in Hubei province, the central part of China, along the Yangzi River. It is leading both of China’s smart cities demonstration project and China’s healthy cities pilot project. -
Chinese Mainland
Address List of Special Warehousing Service Note: The address marked in red are newly added address. (Effective date:October 1, 2021) Province / Directly- controlled City District/county Town, Sub-district and House Number Municipality / Autonomous Region/SAR B4-25, Gate 1, ProLogis Logistics Park, No.1 Tiedi Road, Anhui Province Hefei Shushan District High-tech Zone No.18 Tianzhushan Road, Longshan Sub-district, Wuhu Anhui Province Wuhu Jiujiang District Economic and Technological Development Zone Anhui Province Chuzhou Langya District Longji Leye Photovoltaic Co., Ltd., No.19 Huai'an Road 3/F, No.8 Building, South Area, Lixiang Innovation Park, Anhui Province Chuzhou Nanqiao District Chuzhou, 018 Township Road Anhui Province Chuzhou Nanqiao District No.19 Huai'an Road Yuanrong New Material Holding Co., Ltd., 50 Meters Anhui Province Hefei Shushan District Westward of Bridge of Intersection of Changning Avenue and Ningxi Road Anhui Province Hefei Yaohai District No.88 Dayu Road Anhui Province Hefei Yaohai District No.2177 Dongfang Avenue Beijing BOE Vision-Electronic Technology Co., Ltd., No. Anhui Province Hefei Yaohai District 2177 Dongfang Avenue Anhui Province Hefei Yaohai District No.668 Longzihu Road Anhui Province Hefei Yaohai District No. 668 Longzihu Road Anhui Province Hefei Yaohai District No.2177 Tongling North Road Anhui Province Hefei Yaohai District No.3166 Tongling North Road Anhui Province Hefei Yaohai District No.8 Xiangwang Road Anhui Province Wuhu Jiujiang District No. 8 Anshan Road Anhui Province Wuhu Jiujiang District