Anura: Dendrobatidae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Amphibia: Anura) Da Reserva Catuaba E Seu Entorno, Acre, Amazônia Sul-Ocidental
Efeitos da sucessão florestal sobre a anurofauna (Amphibia: Anura) da Reserva Catuaba e seu entorno, Acre, Amazônia sul-ocidental Vanessa M. de Souza 1; Moisés B. de Souza 2 & Elder F. Morato 2 1 Avenida Assis Chateaubriand 1170, ap. 304, Edifício Azul, Setor Oeste, 74130-011 Goiânia, Goiás, Brasil. E-mail: [email protected] 2 Departamento de Ciências da Natureza, Universidade Federal do Acre. Rodovia BR-364, km 04, Distrito Industrial, 69915-900 Rio Branco, Acre, Brasil. E-mail: [email protected]; [email protected] ABSTRACT. Effect of the forest succession on the anurans (Amphibia(Amphibia: Anura) of the Reserve Catuaba and its peripheryy, Acree, southwestern Amazonia. The objective of this work it was verify the abundance, richness, and the anuran composition in plots of vegetation of different succession stages in a forest and the matrix that surrounds it, of Acre (10º04’S, 67º37’W). The sampling was carried out between August 2005 and April 2006 in twelve plots located in three different sites in the forest. In each site four kinds of environments were chosen: primary forest (wood), secondary forest (capoeira), periphery (matrix) and secondary forest (succession). A total of twenty-seven species distributed in seven families was found. Greater abundance was registered in the plots of matrix two and capoeira three and the least in succession one. The richness was greater in matrix two, with the greater number of exclusive species. The abundance of anurans correlated significantly, with the average circum- ference at the breast height of the trees of the plots. The richness however correlated only marginally, with this structural feature. -

Sexual Maturity and Sexual Dimorphism in a Population of the Rocket-Frog Colostethus Aff
Tolosa et al. Actual Biol Volumen 37 / Número 102, 2014 Sexual maturity and sexual dimorphism in a population of the rocket-frog Colostethus aff. fraterdanieli (Anura: Dendrobatidae) on the northeastern Cordillera Central of Colombia Madurez y dimorfismo sexual de la ranita cohete Colostethus aff. fraterdanieli (Anura: Dendrobatidae) en una población al este de la Cordillera Central de Colombia Yeison Tolosa1, 2, *, Claudia Molina-Zuluaga1, 4,*, Adriana Restrepo1, 5, Juan M. Daza1, ** Abstract The minimum size of sexual maturity and sexual dimorphism are important life history traits useful to study and understand the population dynamics of any species. In this study, we determined the minimum size at sexual maturity and the existence of sexual dimorphism in a population of the rocket-frog, Colostethus aff. fraterdanieli, by means of morphological and morphometric data and macro and microscopic observation of the gonads. Females attained sexual maturity at 17.90 ± 0.1 mm snout-vent length (SVL), while males attained sexual maturity at 16.13 ± 0.06 mm SVL. Females differed from males in size, shape and throat coloration. Males were smaller than females and had a marked and dark throat coloration that sometimes extended to the chest, while females lacked this characteristic, with a throat either immaculate or weakly pigmented. In this study, we describe some important aspects of the reproductive ecology of a population of C. aff. fraterdanieli useful as a baseline for other more specialized studies. Key words: Amphibian, Andes, gonads, histology, morphometry, reproduction Resumen El tamaño mínimo de madurez sexual y el dimorfismo sexual son importantes características de historia de vida, útiles para estudiar y comprender la dinámica poblacional de cualquier especie. -

Amazon Alive: a Decade of Discoveries 1999-2009
Amazon Alive! A decade of discovery 1999-2009 The Amazon is the planet’s largest rainforest and river basin. It supports countless thousands of species, as well as 30 million people. © Brent Stirton / Getty Images / WWF-UK © Brent Stirton / Getty Images The Amazon is the largest rainforest on Earth. It’s famed for its unrivalled biological diversity, with wildlife that includes jaguars, river dolphins, manatees, giant otters, capybaras, harpy eagles, anacondas and piranhas. The many unique habitats in this globally significant region conceal a wealth of hidden species, which scientists continue to discover at an incredible rate. Between 1999 and 2009, at least 1,200 new species of plants and vertebrates have been discovered in the Amazon biome (see page 6 for a map showing the extent of the region that this spans). The new species include 637 plants, 257 fish, 216 amphibians, 55 reptiles, 16 birds and 39 mammals. In addition, thousands of new invertebrate species have been uncovered. Owing to the sheer number of the latter, these are not covered in detail by this report. This report has tried to be comprehensive in its listing of new plants and vertebrates described from the Amazon biome in the last decade. But for the largest groups of life on Earth, such as invertebrates, such lists do not exist – so the number of new species presented here is no doubt an underestimate. Cover image: Ranitomeya benedicta, new poison frog species © Evan Twomey amazon alive! i a decade of discovery 1999-2009 1 Ahmed Djoghlaf, Executive Secretary, Foreword Convention on Biological Diversity The vital importance of the Amazon rainforest is very basic work on the natural history of the well known. -

Etar a Área De Distribuição Geográfica De Anfíbios Na Amazônia
Universidade Federal do Amapá Pró-Reitoria de Pesquisa e Pós-Graduação Programa de Pós-Graduação em Biodiversidade Tropical Mestrado e Doutorado UNIFAP / EMBRAPA-AP / IEPA / CI-Brasil YURI BRENO DA SILVA E SILVA COMO A EXPANSÃO DE HIDRELÉTRICAS, PERDA FLORESTAL E MUDANÇAS CLIMÁTICAS AMEAÇAM A ÁREA DE DISTRIBUIÇÃO DE ANFÍBIOS NA AMAZÔNIA BRASILEIRA MACAPÁ, AP 2017 YURI BRENO DA SILVA E SILVA COMO A EXPANSÃO DE HIDRE LÉTRICAS, PERDA FLORESTAL E MUDANÇAS CLIMÁTICAS AMEAÇAM A ÁREA DE DISTRIBUIÇÃO DE ANFÍBIOS NA AMAZÔNIA BRASILEIRA Dissertação apresentada ao Programa de Pós-Graduação em Biodiversidade Tropical (PPGBIO) da Universidade Federal do Amapá, como requisito parcial à obtenção do título de Mestre em Biodiversidade Tropical. Orientador: Dra. Fernanda Michalski Co-Orientador: Dr. Rafael Loyola MACAPÁ, AP 2017 YURI BRENO DA SILVA E SILVA COMO A EXPANSÃO DE HIDRELÉTRICAS, PERDA FLORESTAL E MUDANÇAS CLIMÁTICAS AMEAÇAM A ÁREA DE DISTRIBUIÇÃO DE ANFÍBIOS NA AMAZÔNIA BRASILEIRA _________________________________________ Dra. Fernanda Michalski Universidade Federal do Amapá (UNIFAP) _________________________________________ Dr. Rafael Loyola Universidade Federal de Goiás (UFG) ____________________________________________ Alexandro Cezar Florentino Universidade Federal do Amapá (UNIFAP) ____________________________________________ Admilson Moreira Torres Instituto de Pesquisas Científicas e Tecnológicas do Estado do Amapá (IEPA) Aprovada em de de , Macapá, AP, Brasil À minha família, meus amigos, meu amor e ao meu pequeno Sebastião. AGRADECIMENTOS Agradeço a CAPES pela conceção de uma bolsa durante os dois anos de mestrado, ao Programa de Pós-Graduação em Biodiversidade Tropical (PPGBio) pelo apoio logístico durante a pesquisa realizada. Obrigado aos professores do PPGBio por todo o conhecimento compartilhado. Agradeço aos Doutores, membros da banca avaliadora, pelas críticas e contribuições construtivas ao trabalho. -

Dietary Resource Use by an Assemblage of Terrestrial Frogs from the Brazilian Cerrado
NORTH-WESTERN JOURNAL OF ZOOLOGY 15 (2): 135-146 ©NWJZ, Oradea, Romania, 2019 Article No.: e181502 http://biozoojournals.ro/nwjz/index.html Dietary resource use by an assemblage of terrestrial frogs from the Brazilian Cerrado Thiago MARQUES-PINTO1,*, André Felipe BARRETO-LIMA1,2,3 and Reuber Albuquerque BRANDÃO1 1. Laboratório de Fauna e Unidades de Conservação, Departamento de Engenharia Florestal, Universidade de Brasília, Brasília – DF, Brazil. 70.910-900, [email protected] 2. Departamento de Ciências Fisiológicas, Instituto de Ciências Biológicas, Universidade de Brasília, Campus Darcy Ribeiro, Brasília – DF, Brazil. 70.910-900, [email protected] 3. Programa de Pós-Graduação em Ecologia, Instituto de Ciências Biológicas, Universidade de Brasília, Campus Darcy Ribeiro, Brasília – DF, Brazil. 70.910-900. *Corresponding author, T. Marques-Pinto, E-mail: [email protected] Received: 08. June 2016 / Accepted: 07. April 2018 / Available online: 19. April 2018 / Printed: December 2019 Abstract. Diet is an important aspect of the ecological niche, and assemblages are often structured based on the ways food resources are partitioned among coexisting species. However, few works investigated the use of food resources in anuran communities in the Brazilian Cerrado biome. Thereby, we studied the feeding ecology of an anuran assembly composed of six terrestrial frog species in a Cerrado protected area. Our main purpose was to detect if there was a structure in the assemblage based on the species’ diet, in terms of the feeding niche overlap and the species’ size. All specimens were collected by pitfall traps placed along a lagoon margin, during the rainy season. We collected six frog species: Elachistocleis cesarii (172 individuals), Leptodactylus fuscus (10), L. -

Volume 2. Animals
AC20 Doc. 8.5 Annex (English only/Seulement en anglais/Únicamente en inglés) REVIEW OF SIGNIFICANT TRADE ANALYSIS OF TRADE TRENDS WITH NOTES ON THE CONSERVATION STATUS OF SELECTED SPECIES Volume 2. Animals Prepared for the CITES Animals Committee, CITES Secretariat by the United Nations Environment Programme World Conservation Monitoring Centre JANUARY 2004 AC20 Doc. 8.5 – p. 3 Prepared and produced by: UNEP World Conservation Monitoring Centre, Cambridge, UK UNEP WORLD CONSERVATION MONITORING CENTRE (UNEP-WCMC) www.unep-wcmc.org The UNEP World Conservation Monitoring Centre is the biodiversity assessment and policy implementation arm of the United Nations Environment Programme, the world’s foremost intergovernmental environmental organisation. UNEP-WCMC aims to help decision-makers recognise the value of biodiversity to people everywhere, and to apply this knowledge to all that they do. The Centre’s challenge is to transform complex data into policy-relevant information, to build tools and systems for analysis and integration, and to support the needs of nations and the international community as they engage in joint programmes of action. UNEP-WCMC provides objective, scientifically rigorous products and services that include ecosystem assessments, support for implementation of environmental agreements, regional and global biodiversity information, research on threats and impacts, and development of future scenarios for the living world. Prepared for: The CITES Secretariat, Geneva A contribution to UNEP - The United Nations Environment Programme Printed by: UNEP World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge CB3 0DL, UK © Copyright: UNEP World Conservation Monitoring Centre/CITES Secretariat The contents of this report do not necessarily reflect the views or policies of UNEP or contributory organisations. -

3 Translation from Norwegian Regulation on the Import
Translation from Norwegian Regulation on the import, export, re-export and transfer or possession of threatened species of wild flora and fauna (Convention on International Trade in Endangered Species, CITES) Commended by Royal Decree of xx xx 2016 on the authority of the Act of 19 June 2009 no. 100 relating to the Management of Nature Diversity, section 26; the Act of 15 June 2001 no. 79 relating to Environmental Protection on Svalbard, section 26, second paragraph: and the Act of 27 February 1930 no. 2 relating to Jan Mayen, section 2, third paragraph. Commended by Ministry of Climate and Environment. Chapter 1 - Purpose and scope 1. Purpose The purpose of this Regulation is to conserve natural wild species which are, or may become, threatened with extinction as the result of trade. 2. Objective scope This Regulation concerns the import, export and re-export of specimens, alive or dead, of animal and plant species cited in Annex 1. Re-export shall mean export of any specimen that has previously been introduced into the Regulation area. This Regulation also concerns domestic transfer and possession of specimens, alive or dead, of animal and plant species cited in Annex 1. The first and second subparagraphs also concern parts of products that are prepared from or declared as prepared from such species. Hunting trophies are also considered to be dead specimens/ products. Hunting trophy means the whole or recognisable parts of animals, either raw, processed or produced. The first, second and third subparagraphs also concern hybrids. Hybrid means the re-crossing of specimens of species regulated under CITES as far back as the fourth generation, with specimens of species not regulated under CITES. -
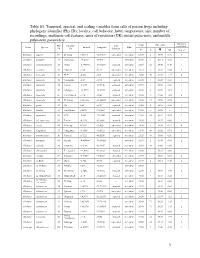
1 Table S1. Temporal, Spectral, and Scaling Variables from Calls Of
Table S1. Temporal, spectral, and scaling variables from calls of poison frogs including phylogeny identifier (Phy ID), locality, call behavior, habit, temperature, size, number of recordings, multinote call features, units of repetition (UR), initial pulse-note, and middle pulse-note parameters. Analyzed Phy Locality Call Temp SVL (mm) Genus Species Latitude Longitude Habit recordings ID ID Behavior °C N ! SD N of ♂ Allobates algorei 60 El Tama 7.65375 -72.19137 concealed terrestrial 23.50 8 18.90 0.70 3 Allobates brunneus 37 Guimaraes -15.2667 -55.5311 -- terrestrial 26.50 1 16.13 0.00 1 Allobates caeruleodactylus 48 Borba -4.398593 -59.60251 exposed terrestrial 25.60 12 15.50 0.40 1 Allobates crombiei 52 Altamira -3.65 -52.38 concealed terrestrial 24.10 2 18.10 0.04 2 Allobates femoralis 43 ECY -0.633 -76.5 concealed terrestrial 25.60 20 23.58 1.27 6 Allobates femoralis 46 Porongaba -8.67 -72.78 exposed terrestrial 25.00 1 25.38 0.00 1 Allobates femoralis 44 Leticia -4.2153 -69.9406 exposed terrestrial 25.50 1 20.90 0.00 1 Allobates femoralis 40 Albergue -12.8773 -71.3865 exposed terrestrial 26.00 6 21.98 2.18 1 Allobates femoralis 41 CAmazonico -12.6 -70.08 exposed terrestrial 26.00 12 22.43 1.06 4 Allobates femoralis 45 El Palmar 8.333333 -61.66667 concealed terrestrial 24.00 27 25.50 0.76 1 Allobates granti 49 FG 3.62 -53.17 exposed terrestrial 24.60 8 16.15 0.55 1 Allobates humilis 59 San Ramon 8.8678 -70.4861 concealed terrestrial 19.50 -- 21.80 -- 1 Allobates insperatus 54 ECY -0.633 -76.4005 exposed terrestrial 24.60 18 16.64 0.93 7 Allobates aff. -

Plan De Manejo Ambiental Reserva Forestal Protectora Tolima Municipio De Gachala Convenio De Cooperacion N° 217 De 2013 Bogot
CONVENIO DE COOPERACION N° 217 DE 2013 SUSCRITO ENTRE CORPOGUAVIO Y LA ONF ANDINA PLAN DE MANEJO AMBIENTAL RESERVA FORESTAL PROTECTORA TOLIMA MUNICIPIO DE GACHALA CONVENIO DE COOPERACION N° 217 DE 2013 BOGOTÁ, DICIEMBRE DE 2013 DIRECCIÓN DE ORDENAMIENTO TERRITORIAL Y ÁREAS PROTEGIDAS ONF ANDINA Tel: +57 (1) 704 15 31 / 755 72 84, Fax: +57 (1) 7557285 www.onfandina.com Carrera 48 No 93-61 Bogotá - Colombia Página 1 de 237 CONVENIO DE COOPERACION N° 217 DE 2013 SUSCRITO ENTRE CORPOGUAVIO Y LA ONF ANDINA CORPORACIÓN AUTÓNOMA REGIONAL DEL GUAVIO CORPOGUAVIO Presidente de la República JUAN MANUEL SANTOS CALDERON Ministra de Ambiente y Desarrollo Sostenible LUZ HELENA SARMIENTO VILLAMIZAR Gobernador de Cundinamarca ALVARO CRUZ VARGAS NIVEL DIRECTIVO Director General OSWALDO JIMÉNEZ DÍAZ Secretario General DIEGO ALBERTO ZULETA GARCIA Subdirectora de Planeación MYRIAM AMPARO ANDRADE HERNANDEZ Subdirector de Gestión Ambiental MARCOS MANUEL URQUIJO COLLAZOS Subdirector Administrativo y Financiero LEONEL ANTONIO CALDERÓN URREA Jefe Oficina de Control Interno INGRID LORENA MEDINA PATARROYO Revisor Fiscal JOSÉ VICENTE SALINAS MARTÍNEZ COMPROMETIDOS POR NATURALEZA DIRECCIÓN DE ORDENAMIENTO TERRITORIAL Y ÁREAS PROTEGIDAS ONF ANDINA Tel: +57 (1) 704 15 31 / 755 72 84, Fax: +57 (1) 7557285 www.onfandina.com Carrera 48 No 93-61 Bogotá - Colombia Página 2 de 237 CONVENIO DE COOPERACION N° 217 DE 2013 SUSCRITO ENTRE CORPOGUAVIO Y LA ONF ANDINA ONF ANDINA Director Ejecutivo Jean-Guénolé Cornet Dirección Ordenamiento Territorial y áreas protegidas Cesar A. -

A Review of Chemical Defense in Poison Frogs (Dendrobatidae): Ecology, Pharmacokinetics, and Autoresistance
Chapter 21 A Review of Chemical Defense in Poison Frogs (Dendrobatidae): Ecology, Pharmacokinetics, and Autoresistance Juan C. Santos , Rebecca D. Tarvin , and Lauren A. O’Connell 21.1 Introduction Chemical defense has evolved multiple times in nearly every major group of life, from snakes and insects to bacteria and plants (Mebs 2002 ). However, among land vertebrates, chemical defenses are restricted to a few monophyletic groups (i.e., clades). Most of these are amphibians and snakes, but a few rare origins (e.g., Pitohui birds) have stimulated research on acquired chemical defenses (Dumbacher et al. 1992 ). Selective pressures that lead to defense are usually associated with an organ- ism’s limited ability to escape predation or conspicuous behaviors and phenotypes that increase detectability by predators (e.g., diurnality or mating calls) (Speed and Ruxton 2005 ). Defended organisms frequently evolve warning signals to advertise their defense, a phenomenon known as aposematism (Mappes et al. 2005 ). Warning signals such as conspicuous coloration unambiguously inform predators that there will be a substantial cost if they proceed with attack or consumption of the defended prey (Mappes et al. 2005 ). However, aposematism is likely more complex than the simple pairing of signal and defense, encompassing a series of traits (i.e., the apose- matic syndrome) that alter morphology, physiology, and behavior (Mappes and J. C. Santos (*) Department of Zoology, Biodiversity Research Centre , University of British Columbia , #4200-6270 University Blvd , Vancouver , BC , Canada , V6T 1Z4 e-mail: [email protected] R. D. Tarvin University of Texas at Austin , 2415 Speedway Stop C0990 , Austin , TX 78712 , USA e-mail: [email protected] L. -

Amphibian Ark Number 43 Keeping Threatened Amphibian Species Afloat June 2018
AArk Newsletter NewsletterNumber 43, June 2018 amphibian ark Number 43 Keeping threatened amphibian species afloat June 2018 In this issue... Reintroduction of the Northern Pool Frog to the UK - Progress Report, April 2018 ............... 2 ® Establishment of a captive breeding program for the Kroombit Tinkerfrog .............................. 4 In situ conservation of the Lemur Leaf Frog through habitat improvement and forest management practices in the Guayacán Rainforest Reserve in Costa Rica .................... 6 Neotropical amphibian biology, management and conservation course .................................. 8 Donation provides for equipment upgrades within the Biogeos Foundation facilities, at the Rescue of Endangered Venezuelan Amphibians program in Venezuela ................... 9 New AArk Conservation Grants program, and call for applications .................................. 10 Amphibian Advocates - José Alfredo Hernández Díaz, Africam Safari, Mexico ........ 11 Amphibian Advocates - Dr. Phil Bishop, Co-Chair IUCN SSC ASG............................... 12 AArk Newsletter - Instructions for authors ...... 13 A private donation helps the Valcheta Frog program in Argentina ...................................... 14 A rich food formula to raise tadpoles in captivity........................................................... 16 Vibicaria Conservation Program: creation of an ex situ model for a rediscovered species in Costa Rica ...................................................... 18 Reproduction of Dendropsophus padreluna at -

A Rapid Biological Assessment of the Upper Palumeu River Watershed (Grensgebergte and Kasikasima) of Southeastern Suriname
Rapid Assessment Program A Rapid Biological Assessment of the Upper Palumeu River Watershed (Grensgebergte and Kasikasima) of Southeastern Suriname Editors: Leeanne E. Alonso and Trond H. Larsen 67 CONSERVATION INTERNATIONAL - SURINAME CONSERVATION INTERNATIONAL GLOBAL WILDLIFE CONSERVATION ANTON DE KOM UNIVERSITY OF SURINAME THE SURINAME FOREST SERVICE (LBB) NATURE CONSERVATION DIVISION (NB) FOUNDATION FOR FOREST MANAGEMENT AND PRODUCTION CONTROL (SBB) SURINAME CONSERVATION FOUNDATION THE HARBERS FAMILY FOUNDATION Rapid Assessment Program A Rapid Biological Assessment of the Upper Palumeu River Watershed RAP (Grensgebergte and Kasikasima) of Southeastern Suriname Bulletin of Biological Assessment 67 Editors: Leeanne E. Alonso and Trond H. Larsen CONSERVATION INTERNATIONAL - SURINAME CONSERVATION INTERNATIONAL GLOBAL WILDLIFE CONSERVATION ANTON DE KOM UNIVERSITY OF SURINAME THE SURINAME FOREST SERVICE (LBB) NATURE CONSERVATION DIVISION (NB) FOUNDATION FOR FOREST MANAGEMENT AND PRODUCTION CONTROL (SBB) SURINAME CONSERVATION FOUNDATION THE HARBERS FAMILY FOUNDATION The RAP Bulletin of Biological Assessment is published by: Conservation International 2011 Crystal Drive, Suite 500 Arlington, VA USA 22202 Tel : +1 703-341-2400 www.conservation.org Cover photos: The RAP team surveyed the Grensgebergte Mountains and Upper Palumeu Watershed, as well as the Middle Palumeu River and Kasikasima Mountains visible here. Freshwater resources originating here are vital for all of Suriname. (T. Larsen) Glass frogs (Hyalinobatrachium cf. taylori) lay their