Reviewing Carbon Spiraling Approach to Understand Organic Matter
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

River Incision Into Bedrock: Mechanics and Relative Efficacy of Plucking, Abrasion and Cavitation
River incision into bedrock: Mechanics and relative efficacy of plucking, abrasion and cavitation Kelin X. Whipple* Department of Earth, Atmospheric, and Planetary Science, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139 Gregory S. Hancock Department of Geology, College of William and Mary, Williamsburg, Virginia 23187 Robert S. Anderson Department of Earth Sciences, University of California, Santa Cruz, California 95064 ABSTRACT long term (Howard et al., 1994). These conditions are commonly met in Improved formulation of bedrock erosion laws requires knowledge mountainous and tectonically active landscapes, and bedrock channels are of the actual processes operative at the bed. We present qualitative field known to dominate steeplands drainage networks (e.g., Wohl, 1993; Mont- evidence from a wide range of settings that the relative efficacy of the gomery et al., 1996; Hovius et al., 1997). As the three-dimensional structure various processes of fluvial erosion (e.g., plucking, abrasion, cavitation, of drainage networks sets much of the form of terrestrial landscapes, it is solution) is a strong function of substrate lithology, and that joint spac- clear that a deep appreciation of mountainous landscapes requires knowl- ing, fractures, and bedding planes exert the most direct control. The edge of the controls on bedrock channel morphology. Moreover, bedrock relative importance of the various processes and the nature of the in- channels play a critical role in the dynamic evolution of mountainous land- terplay between them are inferred from detailed observations of the scapes (Anderson, 1994; Anderson et al., 1994; Howard et al., 1994; Tucker morphology of erosional forms on channel bed and banks, and their and Slingerland, 1996; Sklar and Dietrich, 1998; Whipple and Tucker, spatial distributions. -

STREAMS and RUNNING WATER Hydrologic Cycle •What Happens to Precipitation STREAM •River, Creek, Etc
STREAMS AND RUNNING WATER Hydrologic Cycle •What happens to precipitation STREAM •River, Creek, etc. • Banks, Bed • Confined to a channel –Except in time of flood •Floodplain •Longtitudinal Profile Headwaters, mouth Cross-profile V-shaped SHEETWASH –Unchanneled, thin sheet of flowing water – Results in Sheet Erosion Drainage Basin •Tributary • Divide • Examples: –Mississippi River – Amazon River Drainage Patterns •As seen from above • Dendritic (most common) • Radial • Trellis- in tilted or folded layered rock Factors affecting stream erosion and deposition •Velocity- –Distribution in cross-section •Gradient –Usually decreases downstream – Effect on deposition and erosion •Channel shape and roughness • Discharge- –Volume of water moving past a place in time •Usually given as cubic feet per second (cfs) –Usually increases downstream •Discharge- –Volume of water moving past a place in time •Usually given as cubic feet per second (cfs) –Usually increases downstream American River Discharge •In Sacramento: • Average 3,741 c.f.s. • Usual Range: –Winter 9,000 cfs – Last drought 250 cfs •1986 flood 130,000 cfs Stream Erosion •Hydraulic action • Solution- Dissolves rock • Abrasion (by sand and gravel) –Potholes •Deepens valley by eroding river bed • Lateral erosion of banks on outside of curves Transportation of Sediment •Bed load • Suspended load • Dissolved load Deposition •Bars- –moved in floods – deposited as water slows – Placer Deposits (gold, etc.) – Braided Streams •heavy load of sand and gravel • commonly from glacier outwash streams -
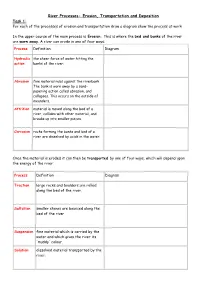
River Processes- Erosion, Transportation and Deposition Task 1: for Each of the Processes of Erosion and Transportation Draw a Diagram Show the Process at Work
River Processes- Erosion, Transportation and Deposition Task 1: For each of the processes of erosion and transportation draw a diagram show the process at work In the upper course of the main process is Erosion. This is where the bed and banks of the river are worn away. A river can erode in one of four ways: Process Definition Diagram Hydraulic the sheer force of water hitting the action banks of the river: Abrasion fine material rubs against the riverbank The bank is worn away by a sand- papering action called abrasion, and collapses. This occurs on the outside of meanders. Attrition material is moved along the bed of a river, collides with other material, and breaks up into smaller pieces. Corrosion rocks forming the banks and bed of a river are dissolved by acids in the water. Once the material is eroded it can then be transported by one of four ways, which will depend upon the energy of the river: Process Definition Diagram Traction large rocks and boulders are rolled along the bed of the river. Saltation smaller stones are bounced along the bed of the river Suspension fine material which is carried by the water and which gives the river its 'muddy' colour. Solution dissolved material transported by the river. In the middle and lower course, the land is much flatter, this means that the river is flowing more slowly and has much less energy. The river starts to deposit (drop) the material that it has been carry Deposition Challenge: Add labels onto the diagram to show where all of the processes could be happening in the river channel. -

Experiments on Patterns of Alluvial Cover and Bedrock Erosion in a Meandering Channel Roberto Fernández1, Gary Parker1,2, Colin P
Earth Surf. Dynam. Discuss., https://doi.org/10.5194/esurf-2019-8 Manuscript under review for journal Earth Surf. Dynam. Discussion started: 27 February 2019 c Author(s) 2019. CC BY 4.0 License. Experiments on patterns of alluvial cover and bedrock erosion in a meandering channel Roberto Fernández1, Gary Parker1,2, Colin P. Stark3 1Ven Te Chow Hydrosystems Laboratory, Department of Civil and Environmental Engineering, University of Illinois at 5 Urbana-Champaign, Urbana, IL 61801, USA 2Department of Geology, University of Illinois at Urbana-Champaign, Urbana, IL 61801, USA 3Lamont Doherty Earth Observatory, Columbia University, Palisades, NY 10964, USA Correspondence to: Roberto Fernández ([email protected]) Abstract. 10 In bedrock rivers, erosion by abrasion is driven by sediment particles that strike bare bedrock while traveling downstream with the flow. If the sediment particles settle and form an alluvial cover, this mode of erosion is impeded by the protection offered by the grains themselves. Channel erosion by abrasion is therefore related to the amount and pattern of alluvial cover, which are functions of sediment load and hydraulic conditions, and which in turn are functions of channel geometry, slope and sinuosity. This study presents the results of alluvial cover experiments conducted in a meandering channel flume of high fixed 15 sinuosity. Maps of quasi-instantaneous alluvial cover were generated from time-lapse imaging of flows under a range of below- capacity bedload conditions. These maps were used to infer patterns -

COLD-BASED GLACIERS in the WESTERN DRY VALLEYS of ANTARCTICA: TERRESTRIAL LANDFORMS and MARTIAN ANALOGS: David R
Lunar and Planetary Science XXXIV (2003) 1245.pdf COLD-BASED GLACIERS IN THE WESTERN DRY VALLEYS OF ANTARCTICA: TERRESTRIAL LANDFORMS AND MARTIAN ANALOGS: David R. Marchant1 and James W. Head2, 1Department of Earth Sciences, Boston University, Boston, MA 02215 [email protected], 2Department of Geological Sciences, Brown University, Providence, RI 02912 Introduction: Basal-ice and surface-ice temperatures are contacts and undisturbed underlying strata are hallmarks of cold- key parameters governing the style of glacial erosion and based glacier deposits [11]. deposition. Temperate glaciers contain basal ice at the pressure- Drop moraines: The term drop moraine is used here to melting point (wet-based) and commonly exhibit extensive areas describe debris ridges that form as supra- and englacial particles of surface melting. Such conditions foster basal plucking and are dropped passively at margins of cold-based glaciers (Fig. 1a abrasion, as well as deposition of thick matrix-supported drift and 1b). Commonly clast supported, the debris is angular and sheets, moraines, and glacio-fluvial outwash. Polar glaciers devoid of fine-grained sediment associated with glacial abrasion include those in which the basal ice remains below the pressure- [10, 12]. In the Dry Valleys, such moraines may be cored by melting point (cold-based) and, in extreme cases like those in glacier ice, owing to the insulating effect of the debris on the the western Dry Valleys region of Antarctica, lack surface underlying glacier. Where cored by ice, moraine crests can melting zones. These conditions inhibit significant glacial exceed the angle of repose. In plan view, drop moraines closely erosion and deposition. -

Avulsion Dynamics in a River with Alternating Bedrock and Alluvial Reaches, Huron River, Northern Ohio (USA)
Open Journal of Modern Hydrology, 2019, 9, 20-39 http://www.scirp.org/journal/ojmh ISSN Online: 2163-0496 ISSN Print: 2163-0461 Avulsion Dynamics in a River with Alternating Bedrock and Alluvial Reaches, Huron River, Northern Ohio (USA) Mark J. Potucek1,2, James E. Evans1 1Department of Geology, Bowling Green State University, Bowling Green, OH, USA 2Arizona Department of Water Resources, Phoenix, AZ, USA How to cite this paper: Potucek, M.J. and Abstract Evans, J.E. (2019) Avulsion Dynamics in a River with Alternating Bedrock and Alluvi- The Huron River consists of alternating bedrock reaches and alluvial reaches. al Reaches, Huron River, Northern Ohio Analysis of historical aerial photography from 1950-2015 reveals six major (USA). Open Journal of Modern Hydrolo- gy, 9, 20-39. channel avulsion events in the 8-km study area. These avulsions occurred in https://doi.org/10.4236/ojmh.2019.91002 the alluvial reaches but were strongly influenced by the properties of the up- stream bedrock reach (“inherited characteristics”). The bedrock reaches Received: December 19, 2018 Accepted: January 20, 2019 aligned with the azimuth of joint sets in the underlying bedrock. One inhe- Published: January 23, 2019 rited characteristic in the alluvial reach downstream is that the avulsion channels diverged only slightly from the orientation of the upstream bedrock Copyright © 2019 by author(s) and channel (range 2˚ - 38˚, mean and standard deviation 12.1˚ ± 13.7˚). A Scientific Research Publishing Inc. This work is licensed under the Creative second inherited characteristic is that avulsion channels were initiated from Commons Attribution International short distances downstream after exiting the upstream bedrock channel reach License (CC BY 4.0). -

The Effects of Longitudinal Differences in Gravel Mobility on the Downstream Fining Pattern in the Cosumnes River, California
The Effects of Longitudinal Differences in Gravel Mobility on the Downstream Fining Pattern in the Cosumnes River, California Candice R. Constantine,1 Jeffrey F. Mount, and Joan L. Florsheim Department of Geological Sciences, University of California, Davis, California 95616, U.S.A. (e-mail: [email protected]) ABSTRACT Downstream fining in the Cosumnes River is partially controlled by longitudinal variation in sediment mobility linked to changes in cross-sectional morphology. Strong fining occurs where the channel is self-formed with section- averaged bankfull dimensionless shear stress (t∗ ) near the threshold of motion (ca. 0.031), allowing for size-selective transport. In contrast, fining is minimal in confined reaches wheret∗ is generally greater than twice the threshold value and transport is nonselective. Downstream fining is best described by a model that depicts fining in discrete intervals separated by a segment in which equal mobility of bed material accounts for the lack of diminishing grain size. Introduction Reduction in the size of bed material with distance 1980; Rice and Church 1998; Rice 1999) or anthro- downstream is commonly observed in gravel-bed pogenic channel modifications (Surian 2002). rivers. Researchers have attributed this feature, The relative importance of sorting and abrasion termed “downstream fining,” to the processes of in determining the diminution coefficient depends selective sorting (Ashworth and Ferguson 1989; on the system in question, particularly on the na- Ferguson and Ashworth 1991; Paola et al. 1992; Fer- ture of material in transport (Bradley 1970; Parker guson et al. 1996; Seal et al. 1997) and abrasion 1991; Werrity 1992; Kodama 1994a, 1994b) and (Bradley 1970; Schumm and Stevens 1973; Werrity whether sediment discharge is limited by transport 1992; Kodama 1994a, 1994b). -

Organic Matter Decomposition and Ecosystem Metabolism As Tools to Assess the Functional Integrity of Streams and Rivers–A Systematic Review
water Review Organic Matter Decomposition and Ecosystem Metabolism as Tools to Assess the Functional Integrity of Streams and Rivers–A Systematic Review Verónica Ferreira 1,* , Arturo Elosegi 2 , Scott D. Tiegs 3 , Daniel von Schiller 4 and Roger Young 5 1 Marine and Environmental Sciences Centre–MARE, Department of Life Sciences, University of Coimbra, 3000–456 Coimbra, Portugal 2 Faculty of Science and Technology, University of the Basque Country (UPV/EHU), P.O. Box 644, 48080 Bilbao, Spain; [email protected] 3 Department of Biological Sciences, Oakland University, Rochester, MI 48309, USA; [email protected] 4 Department of Evolutionary Biology, Ecology and Environmental Sciences, Institut de Recerca de l’Aigua (IdRA), University of Barcelona, Diagonal 643, 08028 Barcelona, Spain; [email protected] 5 Cawthron Institute, Private Bag 2, Nelson 7042, New Zealand; [email protected] * Correspondence: [email protected] Received: 4 November 2020; Accepted: 8 December 2020; Published: 15 December 2020 Abstract: Streams and rivers provide important services to humans, and therefore, their ecological integrity should be a societal goal. Although ecological integrity encompasses structural and functional integrity, stream bioassessment rarely considers ecosystem functioning. Organic matter decomposition and ecosystem metabolism are prime candidate indicators of stream functional integrity, and here we review each of these functions, the methods used for their determination, and their strengths and limitations for bioassessment. We also provide a systematic review of studies that have addressed organic matter decomposition (88 studies) and ecosystem metabolism (50 studies) for stream bioassessment since the year 2000. Most studies were conducted in temperate regions. Bioassessment based on organic matter decomposition mostly used leaf litter in coarse-mesh bags, but fine-mesh bags were also common, and cotton strips and wood were frequent in New Zealand. -

Wind Erosion Abrasion: Effects of Aggregate Moisture
Wind Erosion Abrasion: Effects of Aggregate Moisture L. J. Hagen, E. L. Skidmore, J. B. Layton ASSOC. MEMBER ASAE ABSTRACT One role of moisture in the wind erosion cycle can be FTER rain showers, wind erodible-size particles on described as follows: rain often carries some of the finely Athe surface dry rapidly and often begin abrading the dispersed, water-soluble cementing materials downward, surface aggregates and crust while the latter are still leaving coarse particles such as sand and water-stable moist. The objectives of this investigation were to aggregates at the top. Chepil (1956) studied the effect of measure the abrasive loss rates of moist aggregates and moisture on erosion of these coarse particles by develop prediction equations of the loss rates that would measuring soil loss rates from wind tunnel trays filled be useful in field studies as well as in development of with all erodible particles. He found that the erosion rate complex wind erosion models. Accordingly, soil samples began to decrease whenever soil moisture exceeded 1/3 were obtained from four soils ranging from a sandy loam the moisture content at the —1.5 MPa matric potential. to a silty clay loam. Forty aggregates, 4, to 6 cm in As moisture increased to the moisture content at —1.5 diameter, were selected from each soil and then further MPa, soil loss decreased to nearly zero at wind speeds of subdivided by establishing four moisture levels with 10 14 m/s. Johnson (1965) also reported the effects of both aggregates at each level. Each aggregate was abraded in sand moisture levels and air humidity on threshold wind an enclosed chamber using a calibrated sandblasting speeds of sand. -

Glacial Erosion: Plucking and Abrasion As a Function of Bedrock Properties
CORE Metadata, citation and similar papers at core.ac.uk Provided by NERC Open Research Archive Krabbendam & Glasser: Glacial erosion: Abrasion vs Plucking. MS for Geomorphology Glacial erosion and bedrock properties in NW Scotland: abrasion and plucking, hardness and joint spacing Maarten Krabbendam a,* and Neil F. Glasser b a British Geological Survey, Murchison House, West Mains road, Edinburgh, Scotland, UK b Institute of Geography and Earth Sciences, Aberystwyth University, Aberystwyth, Wales, UK * Corresponding author. Tel: ++44 131 650256 ; E-mail: [email protected] ABSTRACT Subglacial erosion beneath glaciers occurs predominantly by abrasion and plucking, producing distinct erosional forms. The controls on the relative importance of abrasion vs. plucking are poorly understood. On the one hand, glacial conditions that favour or suppress cavity formation (ice velocity, ice thickness, and water pressure) are thought to favour plucking or abrasion, respectively. Conversely, bedrock properties are also known to control landforms, but this has rarely been analysed quantitatively. In this study we compare landforms and bedrock properties of sandstone and quartzite at the bed of a palaeo-ice stream near Ullapool in NW Scotland. The boundary between the rock types is at right angles to the westward palaeo-ice flow, and palaeoglacial conditions on both rock types were similar. We report quantitative parameters for bedrock properties (Schmidt hammer hardness and joint spacing) and use morphometric parameters to analyse the landforms. Torridon sandstone is soft but thick-bedded and with a wide joint spacing. Erosional bedforms include roche moutonnées with smoothed tops and concave stoss sides, whalebacks, and elongate p-forms, indicating a high proportion of abrasion over plucking. -

The Bubble Bursts for Cavitation in Natural Rivers: Laboratory Experiments
The bubble bursts for cavitation in natural rivers: laboratory experiments reveal minor role in bedrock erosion Paul A. Carling1*, Mauricio Perillo2, Jim Best3,4 & Marcelo Garcia4 1College of Environment and Planning, Henan University, Kaifeng, Henan Province, 475004, China, 2 Department of Geology, University of Illinois, Natural History Building, 1301 West Green Street Urbana, IL 61801, 3Departments of Geology, Geography and GIS and Mechanical Science and Engineering, University of Illinois, Natural History Building, 1301 West Green Street Urbana, IL 61801-2938, 4Civil & Environmental Engineering, Ven Te Chow Hydrosystems Laboratory, University of Illinois, 301 N. Mathews Ave., Urbana, IL 61801. *Corresponding author: [email protected]. This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process which may lead to differences between this version and the Version of Record. Please cite this article as doi: 10.1002/esp.4101 This article is protected by copyright. All rights reserved. Abstract The erosion of rock-bedded channels generally is considered a slow process caused mainly by abrasion due to bedload or suspended sediments, but the mechanisms of rapid erosion remain unclear. Cavitation is a clear-fluid erosive process, well known for its effect on engineering structures, when water vapour bubbles collapse and the resultant pressure shocks erode the boundary. However, although the occurrence of cavitation erosion in natural watercourses has long been a matter of debate, as yet there are no incontrovertible examples of cavitation damage to natural river beds. Using flume experiments, we show for the first time that only weakly-cavitating clear- water flows can occur for the range of flow velocities observed in rivers, and these do not erode medium-hardness rocks after 68 hours. -

Sediment Supply Controls Equilibrium Channel Geometry in Gravel Rivers
Sediment supply controls equilibrium channel geometry in gravel rivers Allison M. Pfeiffera,1, Noah J. Finnegana, and Jane K. Willenbringb aDepartment of Earth and Planetary Sciences, University of California, Santa Cruz, CA 95064; and bScripps Institution of Oceanography, University of California, San Diego, CA 92037 Edited by Thomas Dunne, University of California, Santa Barbara, CA, and approved January 26, 2017 (received for review August 3, 2016) τp In many gravel-bedded rivers, floods that fill the channel banks stress ( bf ) may be a criterion to which all gravel rivers must τp create just enough shear stress to move the median-sized gravel conform (8). The critical Shields stress ( c) describes the amount particles on the bed surface (D50). Because this observation is com- of stress needed to initiate median grain motion, normalized for mon and is supported by theory, the coincidence of bankfull flow the grain size, and is generally between 0.03 and 0.08 (9). The and the incipient motion of D has become a commonly used τp 50 bankfull Shields stress ( bf ) describes the stress acting on the bed assumption. However, not all natural gravel channels actually con- during bankfull flow, and (at the reach scale) is approximated as form to this simple relationship; some channels maintain bankfull stresses far in excess of the critical stress required to initiate sed- τp = ρ ðρ ρÞ [1] bf Rbf S s- D50, iment transport. We use a database of >300 gravel-bedded rivers and >600 10Be-derived erosion rates from across North America to 3 where ρ is the density of water (1,000 kg/m ), ρs is the density of explore the hypothesis that sediment supply drives the magnitude sediment, Rbf is the bankfull hydraulic radius, and S is the channel of bankfull shear stress relative to the critical stress required to slope (note that all Shields stresses referred to herein apply to D ).