Organic Matter Decomposition and Ecosystem Metabolism As Tools to Assess the Functional Integrity of Streams and Rivers–A Systematic Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Characterization of Fuel Before and After a Single Prescribed Fire in an Appalachian Hardwood Forest • Elizabeth Bucks, Mary A
Characterization of Fuel before and after a Single Prescribed Fire in an Appalachian Hardwood Forest • Elizabeth bucks, Mary A. Arthur, Jessi E. Lyons, and David L. Loftis Improved understanding of how fuel loads and prescribed fire interact in Appalachian hardwood forests can help managers evaluate the impacts of increased use of prescribed fire in the region. The objective of this study was to characterize fuel loads before and after a single late-winter/early spring prescribed fire and after autumn leaf fall. A repeated measures split-plot design was used to examine dead and down fuels by treatment, sampling time, and landscape position. Preburn mean fuel moss was 40.5 Mg/ha with the duff (Oea) comprising the largest component (19.5 Mg/ha; 48%), followed by large (more than 7.6 cm in diameter) downed lags (9.6 Mg/ha; 24%). Fuel mass was similar across landscape positions; however, duff depth was greater on subxeric compared with intermediate and submesic landscape positions. Burning reduced litter mass (0i; P < 0.00]) and duff depth (P = 0.01). Changes in woody fuels (1-, 10-, 100-, and 1,000-hour) and duff mass were not statistically significant. Post—leaf fall fuel masses did not differ from preburn masses. Thus, a single prescribed burn did not accomplish significant fuel reduction. However, significant declines in duff depth and fuel bed continuity may limit the spread of fire beyond leaf fall and increase potential for soil erosion. This study contributes to the dialogue regarding the use of fire in the Appalachian forest region and impacts on fuel loads. -

Effects of Eutrophication on Stream Ecosystems
EFFECTS OF EUTROPHICATION ON STREAM ECOSYSTEMS Lei Zheng, PhD and Michael J. Paul, PhD Tetra Tech, Inc. Abstract This paper describes the effects of nutrient enrichment on the structure and function of stream ecosystems. It starts with the currently well documented direct effects of nutrient enrichment on algal biomass and the resulting impacts on stream chemistry. The paper continues with an explanation of the less well documented indirect ecological effects of nutrient enrichment on stream structure and function, including effects of excess growth on physical habitat, and alterations to aquatic life community structure from the microbial assemblage to fish and mammals. The paper also dicusses effects on the ecosystem level including changes to productivity, respiration, decomposition, carbon and other geochemical cycles. The paper ends by discussing the significance of these direct and indirect effects of nutrient enrichment on designated uses - especially recreational, aquatic life, and drinking water. 2 1. Introduction 1.1 Stream processes Streams are all flowing natural waters, regardless of size. To understand the processes that influence the pattern and character of streams and reduce natural variation of different streams, several stream classification systems (including ecoregional, fluvial geomorphological, and stream order classification) have been adopted by state and national programs. Ecoregional classification is based on geology, soils, geomorphology, dominant land uses, and natural vegetation (Omernik 1987). Fluvial geomorphological classification explains stream and slope processes through the application of physical principles. Rosgen (1994) classified stream channels in the United States into seven major stream types based on morphological characteristics, including entrenchment, gradient, width/depth ratio, and sinuosity in various land forms. -

Terrestrial Decomposition
Terrestrial Decomposition • Objectives – Controls over decomposition • Litter breakdown • Soil organic matter formation and dynamics – Carbon balance of ecosystems • Soil carbon storage 1 Overview • In terrestrial ecosystems, soils (organic horizon + mineral soil) > C than in vegetation and atmosphere combined 2 Overview • Decomposition is: 1. Major pathway for C loss from ecosystems 2. Central to ecosystem C loss and storage 3 Overview 4 Incorporation 1 year later Overview • Predominant controls on litter decomposition are fairly well constrained 1. Temperature and moisture 2. Litter quality • N availability • Lignin concentration • Lignin:N • Mechanisms for soil organic matter stabilization: 1. Recalcitrance (refers to chemistry) 2. Physical protection • Within soil aggregates • Organo-mineral associations 3. Substrate supply regulation (energetic limitation) 5 Overview • Disturbance can override millenia in a matter of days or years: 1. Land use change 2. Invasive species 3. Climate change • Understanding the mechanistic drivers of decomposition, soil organic matter formation, and carbon stabilization help us make management decisions, take mitigation steps, and protect resources. 6 Overview Native Ōhiʻa - Koa forest Conversion to Reforestation in grass-dominated pasture (80 yr) Eucalyptus plantation (10 yr) Conventional sugar cane harvest. 20° C 18° C 16° C 14° C 7 Sustainable ratoon harvest. Decomposition • Decomposition is the biological, physical and chemical breakdown of organic material – Provides energy for microbial growth -

The Nature and Dynamics of Soil Organic Matter: Plant Inputs, Microbial Transformations, and Organic Matter Stabilization
Soil Biology & Biochemistry 98 (2016) 109e126 Contents lists available at ScienceDirect Soil Biology & Biochemistry journal homepage: www.elsevier.com/locate/soilbio Review paper The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization Eldor A. Paul Natural Resource Ecology Laboratory and Department of Soil and Crop Sciences, Colorado State University, Fort Collins, CO 80523-1499, USA article info abstract Article history: This review covers historical perspectives, the role of plant inputs, and the nature and dynamics of soil Received 19 November 2015 organic matter (SOM), often known as humus. Information on turnover of organic matter components, Received in revised form the role of microbial products, and modeling of SOM, and tracer research should help us to anticipate 31 March 2016 what future research may answer today's challenges. Our globe's most important natural resource is best Accepted 1 April 2016 studied relative to its chemistry, dynamics, matrix interactions, and microbial transformations. Humus has similar, worldwide characteristics, but varies with abiotic controls, soil type, vegetation inputs and composition, and the soil biota. It contains carbohydrates, proteins, lipids, phenol-aromatics, protein- Keywords: Soil organic matter derived and cyclic nitrogenous compounds, and some still unknown compounds. Protection of trans- 13C formed plant residues and microbial products occurs through spatial inaccessibility-resource availability, 14C aggregation of mineral and organic constituents, and interactions with sesquioxides, cations, silts, and Plant residue decomposition clays. Tracers that became available in the mid-20th century made the study of SOM dynamics possible. Soil carbon dynamics Carbon dating identified resistant, often mineral-associated, materials to be thousands of years old, 13 Humus especially at depth in the profile. -

River Incision Into Bedrock: Mechanics and Relative Efficacy of Plucking, Abrasion and Cavitation
River incision into bedrock: Mechanics and relative efficacy of plucking, abrasion and cavitation Kelin X. Whipple* Department of Earth, Atmospheric, and Planetary Science, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139 Gregory S. Hancock Department of Geology, College of William and Mary, Williamsburg, Virginia 23187 Robert S. Anderson Department of Earth Sciences, University of California, Santa Cruz, California 95064 ABSTRACT long term (Howard et al., 1994). These conditions are commonly met in Improved formulation of bedrock erosion laws requires knowledge mountainous and tectonically active landscapes, and bedrock channels are of the actual processes operative at the bed. We present qualitative field known to dominate steeplands drainage networks (e.g., Wohl, 1993; Mont- evidence from a wide range of settings that the relative efficacy of the gomery et al., 1996; Hovius et al., 1997). As the three-dimensional structure various processes of fluvial erosion (e.g., plucking, abrasion, cavitation, of drainage networks sets much of the form of terrestrial landscapes, it is solution) is a strong function of substrate lithology, and that joint spac- clear that a deep appreciation of mountainous landscapes requires knowl- ing, fractures, and bedding planes exert the most direct control. The edge of the controls on bedrock channel morphology. Moreover, bedrock relative importance of the various processes and the nature of the in- channels play a critical role in the dynamic evolution of mountainous land- terplay between them are inferred from detailed observations of the scapes (Anderson, 1994; Anderson et al., 1994; Howard et al., 1994; Tucker morphology of erosional forms on channel bed and banks, and their and Slingerland, 1996; Sklar and Dietrich, 1998; Whipple and Tucker, spatial distributions. -

Plant Species Effects on Nutrient Cycling: Revisiting Litter Feedbacks
Review Plant species effects on nutrient cycling: revisiting litter feedbacks Sarah E. Hobbie Department of Ecology, Evolution and Behavior, University of Minnesota, St. Paul, MN 55108, USA In a review published over two decades ago I asserted reinforce those gradients and patterns of NPP, focusing that, along soil fertility gradients, plant traits change in on feedbacks operating through plant litter decomposition. ways that reinforce patterns of soil fertility and net Specifically, I evaluate two key assumptions underlying primary productivity (NPP). I reevaluate this assertion the plant litter feedback idea: (i) plant litter traits vary in light of recent research, focusing on feedbacks to NPP predictably along fertility gradients, and (ii) such variation operating through litter decomposition. I conclude that reinforces soil fertility gradients through effects on decom- mechanisms emerging since my previous review might position and litter N release. Given the number of synthetic weaken these positive feedbacks, such as negative cross-site analyses of plant traits and their consequences effects of nitrogen on decomposition, while others for nutrient cycling over the past two decades, the time is might strengthen them, such as slower decomposition ripe for revisiting my original assertions. Indeed, I show of roots compared to leaf litter. I further conclude that that my original assertion is more nuanced and complex predictive understanding of plant species effects on than originally claimed. In particular, I discuss the need to nutrient cycling will require developing new frameworks consider leaf litter decomposition more carefully and move that are broadened beyond litter decomposition to con- beyond consideration of leaf litter feedbacks to a more sider the full litter–soil organic matter (SOM) continuum. -

STREAMS and RUNNING WATER Hydrologic Cycle •What Happens to Precipitation STREAM •River, Creek, Etc
STREAMS AND RUNNING WATER Hydrologic Cycle •What happens to precipitation STREAM •River, Creek, etc. • Banks, Bed • Confined to a channel –Except in time of flood •Floodplain •Longtitudinal Profile Headwaters, mouth Cross-profile V-shaped SHEETWASH –Unchanneled, thin sheet of flowing water – Results in Sheet Erosion Drainage Basin •Tributary • Divide • Examples: –Mississippi River – Amazon River Drainage Patterns •As seen from above • Dendritic (most common) • Radial • Trellis- in tilted or folded layered rock Factors affecting stream erosion and deposition •Velocity- –Distribution in cross-section •Gradient –Usually decreases downstream – Effect on deposition and erosion •Channel shape and roughness • Discharge- –Volume of water moving past a place in time •Usually given as cubic feet per second (cfs) –Usually increases downstream •Discharge- –Volume of water moving past a place in time •Usually given as cubic feet per second (cfs) –Usually increases downstream American River Discharge •In Sacramento: • Average 3,741 c.f.s. • Usual Range: –Winter 9,000 cfs – Last drought 250 cfs •1986 flood 130,000 cfs Stream Erosion •Hydraulic action • Solution- Dissolves rock • Abrasion (by sand and gravel) –Potholes •Deepens valley by eroding river bed • Lateral erosion of banks on outside of curves Transportation of Sediment •Bed load • Suspended load • Dissolved load Deposition •Bars- –moved in floods – deposited as water slows – Placer Deposits (gold, etc.) – Braided Streams •heavy load of sand and gravel • commonly from glacier outwash streams -
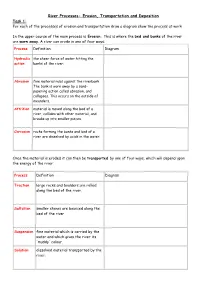
River Processes- Erosion, Transportation and Deposition Task 1: for Each of the Processes of Erosion and Transportation Draw a Diagram Show the Process at Work
River Processes- Erosion, Transportation and Deposition Task 1: For each of the processes of erosion and transportation draw a diagram show the process at work In the upper course of the main process is Erosion. This is where the bed and banks of the river are worn away. A river can erode in one of four ways: Process Definition Diagram Hydraulic the sheer force of water hitting the action banks of the river: Abrasion fine material rubs against the riverbank The bank is worn away by a sand- papering action called abrasion, and collapses. This occurs on the outside of meanders. Attrition material is moved along the bed of a river, collides with other material, and breaks up into smaller pieces. Corrosion rocks forming the banks and bed of a river are dissolved by acids in the water. Once the material is eroded it can then be transported by one of four ways, which will depend upon the energy of the river: Process Definition Diagram Traction large rocks and boulders are rolled along the bed of the river. Saltation smaller stones are bounced along the bed of the river Suspension fine material which is carried by the water and which gives the river its 'muddy' colour. Solution dissolved material transported by the river. In the middle and lower course, the land is much flatter, this means that the river is flowing more slowly and has much less energy. The river starts to deposit (drop) the material that it has been carry Deposition Challenge: Add labels onto the diagram to show where all of the processes could be happening in the river channel. -

The Burdiehouse Burn Valley Park
The Burdiehouse Burn Valley Park Local Nature Reserve Management Plan 2008 – 2018 Revised in 2010 D:\Ranger\My Documents\BBVP\Burdiehouse Burn Valley Park Management Plan 2008\green flag Management plan.doc 1 INDEX PAGE Introduction .................................................................................................................................... 3 SECTION ONE –Site description........................................................................................................... 3 Site maps I, II, III and IV…………………………………………………………………………………………...6-9 1.1 Management plan framework........................................................................................................... 10 SECTION TWO – Our vision ................................................................................................................ 11 SECTION THREE – Aims ..................................................................................................................... 12 3.1 Aims and links with Green Flag criteria ............................................................................................ 12 SECTION FOUR – Surveys .................................................................................................................. 13 4.1 Introduction .................................................................................................................................. 13 4.2 Historical links ................................................................................................................................. -

Organic Matter Decomposition in Simulated Aquaculture Ponds Group Fish Culture and Fisheries Daily Supervisor(S) Dr
O rganic matter decomposition in simulated aquaculture ponds Beatriz Torres Beristain Promotor: Prof. Dr. J.A .J. V erreth H oogleraar in de V isteelt en V isserij W ageningen U niversiteit C o-promotor: Dr. M .C .J. V erdegem U niversitair docent bij the Leerstoelgroep V isteelt en V isserij W ageningen U niversiteit Samenstelling promotiecommissie: Prof. Dr. Y . A vnimelech Technion, Israel Institute of Technology Prof. Dr. Ir. H .J. Gijzen U N ESC O -IH E, Delf, N etherlands Prof. Dr. Ir. M . W .A . V erstegen W ageningen U niversiteit Prof. Dr. Ir. A .A . K oelmans W ageningen U niversiteit Dit onderzoek is uitgevoerd binnen de onderzoekschool W IA S O rganic matter decomposition in simulated aquaculture ponds Beatriz Torres Beristain Proefschrift Ter verkrijging van de graad van doctor O p gezag van de rector magnificus van W ageningen U niversiteit, Prof. Dr. Ir. L. Speelman, In het openbaar te verdedigen O p dinsdag 15 A pril 2005 des namiddags te half tw ee in de A ula Torres Beristain, B. O rganic matter decomposition in simulated aquaculture ponds PhD thesis, Fish C ulture and Fisheries Group, W ageningen Institute of A nimal Sciences. W ageningen U niversity, P.O . Box 338, 6700 A H W ageningen, The N etherlands. - W ith R ef. œW ith summary in Spanish, Dutch and English ISBN : 90-8504-170-8 A Domingo, Y olanda y A lejandro Table of contents C hapter 1 General introduction. 1 C hapter 2 R eview microbial ecology and role in aquaculture ponds. -

Experiments on Patterns of Alluvial Cover and Bedrock Erosion in a Meandering Channel Roberto Fernández1, Gary Parker1,2, Colin P
Earth Surf. Dynam. Discuss., https://doi.org/10.5194/esurf-2019-8 Manuscript under review for journal Earth Surf. Dynam. Discussion started: 27 February 2019 c Author(s) 2019. CC BY 4.0 License. Experiments on patterns of alluvial cover and bedrock erosion in a meandering channel Roberto Fernández1, Gary Parker1,2, Colin P. Stark3 1Ven Te Chow Hydrosystems Laboratory, Department of Civil and Environmental Engineering, University of Illinois at 5 Urbana-Champaign, Urbana, IL 61801, USA 2Department of Geology, University of Illinois at Urbana-Champaign, Urbana, IL 61801, USA 3Lamont Doherty Earth Observatory, Columbia University, Palisades, NY 10964, USA Correspondence to: Roberto Fernández ([email protected]) Abstract. 10 In bedrock rivers, erosion by abrasion is driven by sediment particles that strike bare bedrock while traveling downstream with the flow. If the sediment particles settle and form an alluvial cover, this mode of erosion is impeded by the protection offered by the grains themselves. Channel erosion by abrasion is therefore related to the amount and pattern of alluvial cover, which are functions of sediment load and hydraulic conditions, and which in turn are functions of channel geometry, slope and sinuosity. This study presents the results of alluvial cover experiments conducted in a meandering channel flume of high fixed 15 sinuosity. Maps of quasi-instantaneous alluvial cover were generated from time-lapse imaging of flows under a range of below- capacity bedload conditions. These maps were used to infer patterns -

COLD-BASED GLACIERS in the WESTERN DRY VALLEYS of ANTARCTICA: TERRESTRIAL LANDFORMS and MARTIAN ANALOGS: David R
Lunar and Planetary Science XXXIV (2003) 1245.pdf COLD-BASED GLACIERS IN THE WESTERN DRY VALLEYS OF ANTARCTICA: TERRESTRIAL LANDFORMS AND MARTIAN ANALOGS: David R. Marchant1 and James W. Head2, 1Department of Earth Sciences, Boston University, Boston, MA 02215 [email protected], 2Department of Geological Sciences, Brown University, Providence, RI 02912 Introduction: Basal-ice and surface-ice temperatures are contacts and undisturbed underlying strata are hallmarks of cold- key parameters governing the style of glacial erosion and based glacier deposits [11]. deposition. Temperate glaciers contain basal ice at the pressure- Drop moraines: The term drop moraine is used here to melting point (wet-based) and commonly exhibit extensive areas describe debris ridges that form as supra- and englacial particles of surface melting. Such conditions foster basal plucking and are dropped passively at margins of cold-based glaciers (Fig. 1a abrasion, as well as deposition of thick matrix-supported drift and 1b). Commonly clast supported, the debris is angular and sheets, moraines, and glacio-fluvial outwash. Polar glaciers devoid of fine-grained sediment associated with glacial abrasion include those in which the basal ice remains below the pressure- [10, 12]. In the Dry Valleys, such moraines may be cored by melting point (cold-based) and, in extreme cases like those in glacier ice, owing to the insulating effect of the debris on the the western Dry Valleys region of Antarctica, lack surface underlying glacier. Where cored by ice, moraine crests can melting zones. These conditions inhibit significant glacial exceed the angle of repose. In plan view, drop moraines closely erosion and deposition.