Apoplastic Route Cell Wall Symplastic Route Transmembrane Route Cytosol Key Plasmodesma Plasma Membrane Apoplast Symplast
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Deciphering the Novel Role of Atmin7 in Cuticle Formation and Defense Against the Bacterial Pathogen Infection
International Journal of Molecular Sciences Article Deciphering the Novel Role of AtMIN7 in Cuticle Formation and Defense against the Bacterial Pathogen Infection Zhenzhen Zhao 1, Xianpeng Yang 2 , Shiyou Lü 3, Jiangbo Fan 4, Stephen Opiyo 1, Piao Yang 1 , Jack Mangold 1, David Mackey 5 and Ye Xia 1,* 1 Department of Plant Pathology, College of Food, Agricultural, and Environmental Science, The Ohio State University, Columbus, OH 43210, USA; [email protected] (Z.Z.); [email protected] (S.O.); [email protected] (P.Y.); [email protected] (J.M.) 2 College of Life Sciences, Shandong Normal University, Jinan 250014, China; [email protected] 3 State Key Laboratory of Biocatalysis and Enzyme Engineering, School of Life Sciences, Hubei University, Wuhan 434200, China; [email protected] 4 School of Agriculture and Biology, Shanghai Jiao Tong University, Shanghai 200240, China; [email protected] 5 Department of Horticulture and Crop Science, College of Food, Agricultural, and Environmental Science, The Ohio State University, Columbus, OH 43210, USA; [email protected] * Correspondence: [email protected] Received: 15 July 2020; Accepted: 1 August 2020; Published: 3 August 2020 Abstract: The cuticle is the outermost layer of plant aerial tissue that interacts with the environment and protects plants against water loss and various biotic and abiotic stresses. ADP ribosylation factor guanine nucleotide exchange factor proteins (ARF-GEFs) are key components of the vesicle trafficking system. Our study discovers that AtMIN7, an Arabidopsis ARF-GEF, is critical for cuticle formation and related leaf surface defense against the bacterial pathogen Pseudomonas syringae pathovar tomato (Pto). -

Diversity of Plant Virus Movement Proteins: What Do They Have in Common?
processes Review Diversity of Plant Virus Movement Proteins: What Do They Have in Common? Yuri L. Dorokhov 1,2,* , Ekaterina V. Sheshukova 1, Tatiana E. Byalik 3 and Tatiana V. Komarova 1,2 1 Vavilov Institute of General Genetics Russian Academy of Sciences, 119991 Moscow, Russia; [email protected] (E.V.S.); [email protected] (T.V.K.) 2 Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, 119991 Moscow, Russia 3 Department of Oncology, I.M. Sechenov First Moscow State Medical University, 119991 Moscow, Russia; [email protected] * Correspondence: [email protected] Received: 11 November 2020; Accepted: 24 November 2020; Published: 26 November 2020 Abstract: The modern view of the mechanism of intercellular movement of viruses is based largely on data from the study of the tobacco mosaic virus (TMV) 30-kDa movement protein (MP). The discovered properties and abilities of TMV MP, namely, (a) in vitro binding of single-stranded RNA in a non-sequence-specific manner, (b) participation in the intracellular trafficking of genomic RNA to the plasmodesmata (Pd), and (c) localization in Pd and enhancement of Pd permeability, have been used as a reference in the search and analysis of candidate proteins from other plant viruses. Nevertheless, although almost four decades have passed since the introduction of the term “movement protein” into scientific circulation, the mechanism underlying its function remains unclear. It is unclear why, despite the absence of homology, different MPs are able to functionally replace each other in trans-complementation tests. Here, we consider the complexity and contradictions of the approaches for assessment of the ability of plant viral proteins to perform their movement function. -

Book of Abstracts
CHALLENGES FOR PLANT NUTRITION IN CHANGING ENVIRONMENTS International Workshop and Meeting of the German Society of Plant Nutrition 2012 University of Bonn September 5 – 8, 2012 Book of Abstracts Table of Contents Plenary Session: Introductory talks ................................................. 2 Plenary Session S1: Processes on leaf surfaces ................................. 5 Plenary Session S2: Plant water relations .......................................... 14 Plenary Session S3: Nutrient dynamics in changing environments .... 26 Plenary Session S4: Crop responses to nutrient imbalances ............. 38 Plenary Session S5: Phenotyping and early stress responses ........... 80 Poster Session P1: Fertilization (inorganic) ...................................... 92 Poster Session P2: Fertilization (organic) / Soil amendments ........ 110 Poster Session P3: Nutrient efficiency / Genomics ......................... 131 Poster Session P4: Root physiology / Root-soil interactions........... 148 Poster Session P5: Physiological response to abiotic stress .......... 169 Poster Session P6: Physiological response to nutrient imbalances 182 Poster Session P7: Nutrients and ecosystems / Climate change ... 200 Poster Session P8: Signalling / Quality / Phenotyping .................... 215 1 DGP Meeting September 5-9, 2012 Plenary Session: Introductory talks 2 DGP Meeting September 5-9, 2012 Plant nutrition in a changing environment. Patrick H. Brown Department of Plant Sciences, University of California, Davis, CA 95616, US; E-mail: [email protected] The scientific discipline of plant nutrition is wonderfully broad in its scale and its scope. From the exploration of the function of nutrients as signals and regulators of plant function, to the role of plant nutrients in agricultural productivity and food quality, to the exploration of the effects of nutrient losses on global environments, plant nutrition is a truly integrative discipline and will play a critical role in mans’ ability to adapt to environmental change. -
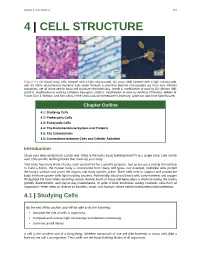
Chapter 4 – Cell Structure
Chapter 4 | Cell Structure 107 4 | CELL STRUCTURE Figure 4.1 (a) Nasal sinus cells (viewed with a light microscope), (b) onion cells (viewed with a light microscope), and (c) Vibrio tasmaniensis bacterial cells (seen through a scanning electron microscope) are from very different organisms, yet all share certain basic cell structure characteristics. (credit a: modification of work by Ed Uthman, MD; credit b: modification of work by Umberto Salvagnin; credit c: modification of work by Anthony D'Onofrio, William H. Fowle, Eric J. Stewart, and Kim Lewis of the Lewis Lab at Northeastern University; scale-bar data from Matt Russell) Chapter Outline 4.1: Studying Cells 4.2: Prokaryotic Cells 4.3: Eukaryotic Cells 4.4: The Endomembrane System and Proteins 4.5: The Cytoskeleton 4.6: Connections between Cells and Cellular Activities Introduction Close your eyes and picture a brick wall. What is the wall's basic building block? It is a single brick. Like a brick wall, cells are the building blocks that make up your body. Your body has many kinds of cells, each specialized for a specific purpose. Just as we use a variety of materials to build a home, the human body is constructed from many cell types. For example, epithelial cells protect the body's surface and cover the organs and body cavities within. Bone cells help to support and protect the body. Immune system cells fight invading bacteria. Additionally, blood and blood cells carry nutrients and oxygen throughout the body while removing carbon dioxide. Each of these cell types plays a vital role during the body's growth, development, and day-to-day maintenance. -

Development and Function of Plasmodesmata in Zygotes of Fucus Distichus
Botanica Marina 2015; 58(3): 229–238 Chikako Nagasato*, Makoto Terauchi, Atsuko Tanaka and Taizo Motomura Development and function of plasmodesmata in zygotes of Fucus distichus Abstract: Brown algae have plasmodesmata, tiny tubu- Introduction lar cytoplasmic channels connecting adjacent cells. The lumen of plasmodesmata is 10–20 nm wide, and it takes a Multicellular organisms such as animals, fungi, land simple form, without a desmotubule (the inner membrane plants, and brown algae have specific cellular connections. structure consisting of endoplasmic reticulum in the plas- These structures connect the cytoplasm of adjacent cells modesmata of green plants). In this study, we analyzed and provide a route for cell-cell communication. Green the ultrastructure and distribution of plasmodesmata plants, including land plants and certain species of green during development of Fucus distichus zygotes. The first algae (reviewed in Robards and Lucas 1990, Raven 1997), cytokinesis of zygotes in brown algae is not accompanied and brown algae (Bisalputra 1966) possess plasmodes- by plasmodesmata formation. As the germlings develop, mata. These are cytoplasmic canals that pass through the plasmodesmata are found in all septal cell walls, includ- septal cell wall and connect adjacent cells. Plasmodes- ing the first cell division plane. Plasmodesmata are formed mata in green plants and brown algae share similar char- de novo on the existing cell wall. Pit fields, which are clus- acteristics; however, plasmodesmata in brown algae lack ters of plasmodesmata, were observed in germlings with a desmotubule, a tubular strand of connecting endoplas- differentiated cell layers. Apart from the normal plas- mic reticulum (ER) that penetrates the plasmodesmata modesmata, these pit fields had branched plasmodes- of land plants (Terauchi et al. -

Testing the Mьnch Hypothesis of Long Distance Phloem Transport in Plants
Downloaded from orbit.dtu.dk on: Oct 10, 2021 Testing the Münch hypothesis of long distance phloem transport in plants Knoblauch, Michael; Knoblauch, Jan; Mullendore, Daniel L.; Savage, Jessica A.; Babst, Benjamin A.; Beecher, Sierra D.; Dodgen, Adam C.; Jensen, Kaare Hartvig; Holbrook, N. Michele Published in: eLife Link to article, DOI: 10.7554/eLife.15341 Publication date: 2016 Document Version Publisher's PDF, also known as Version of record Link back to DTU Orbit Citation (APA): Knoblauch, M., Knoblauch, J., Mullendore, D. L., Savage, J. A., Babst, B. A., Beecher, S. D., Dodgen, A. C., Jensen, K. H., & Holbrook, N. M. (2016). Testing the Münch hypothesis of long distance phloem transport in plants. eLife, 5. https://doi.org/10.7554/eLife.15341 General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. Users may download and print one copy of any publication from the public portal for the purpose of private study or research. You may not further distribute the material or use it for any profit-making activity or commercial gain You may freely distribute the URL identifying the publication in the public portal If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim. RESEARCH ARTICLE Testing -

PLANT PHYSIOLOGY and ANATOMY in RELATION to HERBICIDE ACTION Physiology. James E. Hill Extension Weed Scientist As We Advance To
16 PLANT PHYSIOLOGY AND ANATOMY IN RELATION TO HERBICIDE ACTION James E. Hill Extension Weed Scientist Physiology. As we advance towards herbicides with greater selectivity and more plant toxicity, we will be reouired to know more about plant physiology and anatomy. All too often principles of plant physiology are dismissed as being too complicated to have any practical bearing on herbicide use. Yet many practices regular ly used in the field to obtain proper herbicide selectivity, have their basis of selectivity in the physiology of the plant. Plant anatomy and plant physiology will be considered together in this discussion because plant structure and function are delicately interwoven in the living plant. Plants react to herbicides within the nonnal framework of their anatomy and physiology. There are no plant processes and no structures specifically for herbicides. In fact, the lethal effects of different groups of herbicides are caused by an interference with one or more natural physiological processes in the plant. A convenient way to look at herbicides as related to plant structure and function is to divide the physiological processes into three: 1) absorption, 2) translocation, and 3) site of action. The term absorption simply means uptake, or how a chemical gets into the plant. The term translocation means movement, how a chemical moves from the place where it is absorbed to the place where it will exhibit its legal activity. Lastly, the site of action refers to the process or location where the herbicide reacts to injure or kill the plant. Each of these physiological processes are examined below in relation to herbicide selectivity, the theme of the 1976 Weed School. -

Genome-Wide Identification and Characterization of Apple P3A-Type Atpase Genes, with Implications for Alkaline Stress Responses
Article Genome-Wide Identification and Characterization of Apple P3A-Type ATPase Genes, with Implications for Alkaline Stress Responses Baiquan Ma y , Meng Gao y, Lihua Zhang, Haiyan Zhao, Lingcheng Zhu, Jing Su, Cuiying Li, Mingjun Li , Fengwang Ma * and Yangyang Yuan * State Key Laboratory of Crop Stress Biology for Arid Areas/Shaanxi Key Laboratory of Apple, College of Horticulture, Northwest A&F University, Yangling 712100, China; [email protected] (B.M.); [email protected] (M.G.); [email protected] (L.Z.); [email protected] (H.Z.); [email protected] (L.Z.); [email protected] (J.S.); [email protected] (C.L.); [email protected] (M.L.) * Correspondence: [email protected] (F.M.); [email protected] (Y.Y.); Tel.: +86-029-8708-2648 (F.M.) These authors contributed equally to this work. y Received: 4 January 2020; Accepted: 5 March 2020; Published: 6 March 2020 Abstract: The P3A-type ATPases play crucial roles in various physiological processes via the generation + of a transmembrane H gradient (DpH). However, the P3A-type ATPase superfamily in apple remains relatively uncharacterized. In this study, 15 apple P3A-type ATPase genes were identified based on the new GDDH13 draft genome sequence. The exon-intron organization of these genes, the physical and chemical properties, and conserved motifs of the encoded enzymes were investigated. Analyses of the chromosome localization and ! values of the apple P3A-type ATPase genes revealed the duplicated genes were influenced by purifying selection pressure. Six clades and frequent old duplication events were detected. Moreover, the significance of differences in the evolutionary rates of the P3A-type ATPase genes were revealed. -

PIN-Pointing the Molecular Basis of Auxin Transport Klaus Palme* and Leo Gälweiler†
375 PIN-pointing the molecular basis of auxin transport Klaus Palme* and Leo Gälweiler† Significant advances in the genetic dissection of the auxin signals — signals that co-ordinate plant growth and devel- transport pathway have recently been made. Particularly opment, rather than signals that carry information from relevant is the molecular analysis of mutants impaired in auxin source cells to specific target cells or tissues [13]. Applying transport and the subsequent cloning of genes encoding this conceptual framework to interpret the activities of candidate proteins for the elusive auxin efflux carrier. These plant growth substances such as auxin led to the sugges- studies are thought to pave the way to the detailed tion that auxin might be better viewed as a substance understanding of the molecular basis of several important that — similarly to signals acting in the animal nervous sys- facets of auxin action. tem — collates information from various sources and transmits processed information to target tissues [14]. Addresses Max-Delbrück-Laboratorium in der Max-Planck-Gesellschaft, Carl-von- But if auxin does not act like an animal hormone, how can Linné-Weg 10, D-50829 Köln, Germany we explain its numerous activities? How can we explain, *[email protected] for example, that auxin can act as a mitogen to promote cell †[email protected] division, whereas at another time its action may be better Current Opinion in Plant Biology 1999, 2:375–381 interpreted as a morphogen [15]? The observation that 1369-5266/99/$ — see front matter © 1999 Elsevier Science Ltd. auxin replaces all the correlative effects of a shoot apex led All rights reserved. -

Tracing Root Permeability: Comparison of Tracer Methods
DOI: 10.1007/s10535-016-0634-2 BIOLOGIA PLANTARUM 60 (4): 695-705, 2016 Tracing root permeability: comparison of tracer methods E. PECKOVÁ, E. TYLOVÁ, and A. SOUKUP* Department of Experimental Plant Biology, Faculty of Natural Sciences, Charles University in Prague, CZ-12844 Prague, Czech Republic Abstract Root epidermis and apoplastic barriers (endodermis and exodermis) are the critical root structures involved in setting up plant-soil interface by regulating free apoplastic movement of solutes within root tissues. Probing root apoplast permeability with “apoplastic tracers” presents one of scarce tools available for detection of “apoplastic leakage” sites and evaluation of their role in overall root uptake of water, nutrients, or pollutants. Although the tracers are used for many decades, there is still not an ideal apoplastic tracer and flawless procedure with straightforward interpretation. In this article, we present our experience with the most frequently used tracers representing various types of chemicals with different characteristics. We examine their behaviour, characteristics, and limitations. Here, we show that results gained with an apoplastic tracer assay technique are reliable but depend on many parameters – chemical properties of a selected tracer, plant species, cell wall properties, exposure time, or sample processing. Additional key words: apoplast, berberine, endodermis, exodermis, ferrous ions, PAS reaction, propidium iodide, PTS. Introduction Root permeability is one of the key features determining et al. 2014). root-soil communication, resources acquisition, or Apoplast permeability modulates root uptake resistance to pollutants with implication to plant stress characteristics substantially, but there is a limited set of tolerance or food quality. Passive non-selective transport methodological tools to evaluate its extent and spatial via apoplast is restricted by apoplastic barriers. -

Uncovering Ph at Both Sides of the Root Plasma Membrane Interface Using Noninvasive Imaging
Uncovering pH at both sides of the root plasma membrane interface using noninvasive imaging Alexandre Martinièrea, Rémy Gibrata, Hervé Sentenaca, Xavier Dumonta, Isabelle Gaillarda, and Nadine Parisa,1 aBiochimie et Physiologie Moléculaire des Plantes, Université de Montpellier, Centre National de Recherche Scientifique, L’Institut National de la Recherche Agronomique, Montpellier SupAgro, Université de Montpellier, Montpellier, France Deborah J. Delmer, Emeritus University of California, Davis, CA, and approved April 16, 2018 (received for review December 15, 2017) Building a proton gradient across a biological membrane and between One of the physiological parameters tightly regulated in the different tissues is a matter of great importance for plant development interstitial fluid is pH. For plants, pH regulation is crucial for and nutrition. To gain a better understanding of proton distribution in mineral nutrition, since it participates in the plasma membrane the plant root apoplast as well as across the plasma membrane, we (PM) proton-motive force (PMF), along with the transmembrane generated Arabidopsis plants expressing stable membrane-anchored pH gradient (PM delta pH) and an electrical component. The + ratiometric fluorescent sensors based on pHluorin. These sensors en- PMF is formed by the activity of a P-type H -ATPase and pro- abled noninvasive pH-specific measurements in mature root cells from vides the driving force for the uptake of minerals by transporters – the medium epidermis interface up to the inner cell layers that lie (symporters and antiporters) and channels (9). While the electrical beyond the Casparian strip. The membrane-associated apoplastic component of the PMF can be studied by electrophysiological pH was much more alkaline than the overall apoplastic space pH. -

Plasmodesmata and the Control of Symplastic Transport
Blackwell Science, LtdOxford, UKPCEPlant, Cell and Environment0016-8025Blackwell Publishing Ltd 2002 26 Original Article Plant, Cell and Environment (2003) 26, 103–124 Plasmodesmata and the control of symplastic transport A. G. ROBERTS & K. J. OPARKA Scottish Crop Research Institute, Invergowrie, Dundee, DD2 5DA, UK ‘There are holes in the sky other cells, some degree of intercellular connection is main- where the rain gets in, tained. This plasmodesmal continuum that potentially but they’re ever so small exists throughout the whole plant is termed the symplast that’s why rain’s so thin’ (Münch 1930). However, the symplast is not the open con- Spike Milligan (1968) tinuum that Münch originally hypothesized, but is divided into functional domains, each tightly regulated by different In 1879 Eduard Tangle discovered cytoplasmic connections forms of plasmodesmata (Erwee & Goodwin 1985; Ehlers between cells in the cotyledons of Strychnos nuxvomica, & Kollmann 2001). Plasmodesmata are now thought of as which he interpreted to be protoplasmic contacts. This led fluid, dynamic structures that can be modified both struc- him to hypothesize that ‘the protoplasmic bodies . are turally and functionally to cope with the requirements of united by thin strands passing through connecting ducts in specific cells and tissues. the walls, which put the cells into connection with each other and so unite them to an entity of higher order’ (Carr 1976). This challenged the then current view that cells func- THE STRUCTURE OF PLASMODESMATA tioned as autonomous units. It was after much research in Based on structure, two basic types of plasmodesmata have many other species and cell types that Strasburger, in 1901, been characterized; simple and branched.