The Short-Term Impacts of a Cyclone on Seagrass Communities In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Disaster Relief Emergency Fund (DREF) Madagascar: Tropical Cyclone Haruna
Disaster relief emergency fund (DREF) Madagascar: Tropical cyclone Haruna DREF operation n° MDRMG009 GLIDE n° TC-2013-000022-MDG 01 March, 2013 The International Federation of Red Cross and Red Crescent (IFRC) Disaster Relief Emergency Fund (DREF) is a source of un-earmarked money created by the Federation in 1985 to ensure that immediate financial support is available for Red Cross and Red Crescent emergency response. The DREF is a vital part of the International Federation’s disaster response system and increases the ability of National Societies to respond to disasters. CHF 298,747 has been allocated from the IFRC’s Disaster Relief Emergency Fund (DREF) to support Madagascar Red Cross Society in delivering immediate assistance to some 10,000 beneficiaries. Unearmarked funds to repay DREF are encouraged. Summary: Tropical cyclone Haruna brought heavy rains to the west coast of Madagascar for several days before making landfall on 22 February, 2013. Information from the National Office for Disaster Management, Ministry of Interior (BNGRC) indicates that as of 25 February, Morombe and Toliara are the most affected while Sakaraha, Miandrivazo and MRCS volunteers helped to evacuate families stranded by the floods due to the cyclone. Photo: MRCS Antananarivo are moderately affected. To date, BNGRC has registered 23 deaths, with 16 people missing, 81 people injured and 22,498 others affected, with the numbers increasing every day. Malagasy Red Cross Society (MRCS) activated its contingency plan and mobilized its national disaster response team (NDRT) to assist affected families. Volunteers from its branches assisted with evacuations in the days leading up to the cyclone making landfall, and provided psychosocial support as well as hygiene awareness in the aftermath. -

The Climate in Africa: 2013
The Climate in Africa: 2013 WMO-No. 1147 WMO-No. 1147 © World Meteorological Organization, 2015 The right of publication in print, electronic and any other form and in any language is reserved by WMO. Short extracts from WMO publications may be reproduced without authorization, provided that the complete source is clearly indicated. Editorial correspondence and requests to publish, reproduce or translate this publication in part or in whole should be addressed to: Chairperson, Publications Board World Meteorological Organization (WMO) 7 bis, avenue de la Paix Tel.: +41 (0) 22 730 84 03 P.O. Box 2300 Fax: +41 (0) 22 730 80 40 CH-1211 Geneva 2, Switzerland E-mail: [email protected] ISBN 978-92-63-11147-0 Cover illustration: xxx NOTE The designations employed in WMO publications and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of WMO concerning the legal status of any country, territory, city or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products does not imply that they are endorsed or recommended by WMO in preference to others of a similar nature which are not mentioned or advertised. The findings, interpretations and conclusions expressed in WMO publications with named authors are those of the authors alone and do not necessarily reflect those of WMO or its Members. Contents Foreword . 3 Climate assessment: key continental features . 4 Temperature 4 Precipitation 5 Climate assessment: subregional features. 7 Southern Africa . .7 . Temperature 7 Precipitation 7 South-West Indian Ocean Island Countries . -

DREF Final Report Madagascar: Tropical Cyclone Haruna
DREF Final Report Madagascar: Tropical cyclone Haruna DREF operation n° MDRMG009 GLIDE n° TC-2013-000022-MDG 20 December, 2013 The International Federation of Red Cross and Red Crescent (IFRC) Disaster Relief Emergency Fund (DREF) is a source of un-earmarked money created by the Federation in 1985 to ensure that immediate financial support is available for Red Cross and Red Crescent emergency response. The DREF is a vital part of the International Federation’s disaster response system and increases the ability of National Societies to respond to disasters. Period covered by this Final Report: 28 February to 31 August, 2013. Summary: CHF 298,747 was allocated from the IFRC’s Disaster Relief Emergency Fund (DREF) on 28 February 2013 to support the National Society in delivering assistance to some 10,000 beneficiaries (2,000 vulnerable households) affected by the Tropical cyclone Haruna. This DREF was initially allocated on 28 February 2013 for CHF 298,747 for three months to assist 10,000 beneficiaries. An operations update n°1 was issued on 7 May, 2013 to revise the planned activities after in-depth assessments as well as Malagasy RC providing potable water in Ankatsakatsa extend the operation time frame by a month. Sakaraha. Photo/ Malagasy RC Operations update n°2 was issued 1 October, 2013 further extending the operation timeframe to the end of August 2013. The DREF operation has enabled the Malagasy RC (MRCS) to respond to the emergency needs of 2,000 families affected by the disaster through the distribution of shelter and non-food items, potable water and health and hygiene promotion activities. -

Regional Inter-Agency Standing Committee (RIASCO), Southern Africa
Humanitarian Trends in Southern Africa: Challenges and Opportunities Regional Inter-Agency Standing Committeei (RIASCO), Southern Africa Humanitarian Trends in Southern Africa: Challenges and Opportunities FOR FURTHER INFORMATION, PLEASE CONTACT Disaster Mitigation for Sustainable Livelihoods Programme Stellenbosch University Private Bag x1 Matieland, 7602 South Africa [email protected] PLEASE CITE AS: Holloway A., Chasi V., de Waal J., Drimie S., Fortune G., Mafuleka G., Morojele M., Penicela Nhambiu B., Randrianalijaona M., Vogel C. and Zweig P. 2013. Humanitarian Trends in Southern Africa: Challenges and Opportunities. Regional Interagency Standing Committee, Southern Africa. Rome, FAO. COVER IMAGE CAPTION “PRECIOUS BUNDLE: An as-yet-unnamed baby who was born on the back of a bakkie (pick- up truck) just days ago during the deluge is carried gently across the water by a farmworker” – courtesy of Antione de Ras, The Star, January 25, 2013. i The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. The views expressed in this information product are those of the author(s) and do not necessarily reflect the views or policies of FAO. -

Humanitarian Bulletin
Humanitarian Bulletin Southern Africa Issue 17 | December 2014 In this issue Issue 00 | DD – DD Mmm YYYY The importance of being prepared P. 1 Current levels of preparedness P. 2 HIGHLIGHTS Preparedness case studies P. 4 The benefits of strengthening disaster preparedness are cost effectiveness and the delivery of SADC, IGES/COLA effective humanitarian response. Countries in the region have The importance of being prepared varying levels of preparedness. Mozambique and Madagascar Spending to save are most exposed to tropical Preparedness comprises activities that contribute to the pre-planned, timely and effective cyclones. response of individuals and communities to reduce the impact of a hazard and its consequences. Moreover, investing in preparedness makes sense. The average global FIGURES* economic cost of disasters increased approximately six-fold from 1970 to 2000. The Year People affected by World Bank and US Geological Survey suggest that investments of US$40 billion in storms/floods preparedness, prevention and mitigation would have reduced global economic losses 2007/8 1,050,000 caused by disasters in the 1990s by $280 billion. According to UNDP and OCHA, every $1 invested in prevention could save $7 in recovery. 2008/9 1,370,000 2009/10 367,000 As we enter the Southern Africa rainfall season, which stretches from October to April, 2010/11 708,000 this bulletin will provide an overview of current levels of flood preparedness in the region. 2011/12 554,000 2012/13 519,000 Floods: a cyclical disaster 2013/14 453,256 Every year hundreds of thousands of people are affected by flooding in Southern Africa. -

(Ssip) 2019 Geography
SECONDARY SCHOOL IMPROVEMENT PROGRAMME (SSIP) 2019 GEOGRAPHY REVISED LEARNER NOTES SESSION 1: CLIMATOLOGY GRADE 12 G E O G R A P H Y - F I R S T P A P E R How to achieve 30% Add this to achieve 60% Add this to achieve 70%+ MIDLATITUDE CYCLONES ADD THE FOLLOWING ADD THE FOLLOWING General characteristics MIDLATITUDE CYCLONES MIDLATITUDE CYCLONES Cold front weather changes Areas where formed Conditions for formation C Identify stages and reasons Weather associated with cold Weather associated with cold, TROPICAL CYCLONES and warm fronts warm and occluded fronts L General characteristics TROPICAL CYCLONES Cross sections: cold, warm and I Identify stages and reasons Areas where formed occluded fronts M SUBTROPICAL ANTICYCLONES Conditions for formation TROPICAL CYCLONES A 3 high pressure cells (location) SUB-TROPICAL ANTI- Associated weather patterns T Formation: line thunderstorms CYCLONES SUB-TROPICAL ANTI- South African berg winds Influence-high pressure cells CYCLONES E VALLEY CLIMATES VALLEY CLIMATES Coastal low pressures Aspect (which slope is warmer) Frost pockets radiation fog Anabatic/katabatic winds Influence on farming and Inversions settlements URBAN CLIMATES URBAN CLIMATES Why are cities warmer? Strategies to reduce heat islands Definitions: heat island and pollution domes DRAINAGE SYSTEMS ADD THE FOLLOWING ADD THE FOLLOWING G ALL concepts DRAINAGE SYSTEMS FLUVIAL PROCESSES E Drainage patterns (all) Types of rivers River grading Laminar and turbulent flow Drainage density (high/low) Superimposed -
Madagascar Sitrep 2 TC Haruna 23 Feb 2013
Madagascar: Tropical Cyclone Haruna Office of the Resident Coordinator Situation Report No. 2 (as of 23 Feb 2013) This report is produced by Office of the Resident Coordinator in Madagascar in collaboration with humanitarian partners. It covers the period from 22 to 23 February 2013. The next report will be issued around 25 February 2013. Highlights Tropical Cyclone Haruna made landfall over the south-west coast of Madagascar on 22 February 2013 as a powerful Category 2 Tropical Cyclone. Haruna made its exit to the Indian Ocean, as a tropical storm, around Taolagnaro at about 09:00 on 23 February. The most affected districts are Morombe and Toliary. Sakaraha and Betioky Sud are moderately affected. On 23 February, Fiherenana dyke burst, causing flooding to a large area and 25,000 people potentially needed to be evacuated. To date, some 1,000 people have been evacuated to temporary sites and 5,000 others are being hosted by their relatives. Initial situation assessments by local authorities and humanitarian partners partially started on 23 February. Reports of significant damage to houses, public buildings and infrastructure are being received. Aerial assessments will be conducted on 24 and 25 February by the Government, INGOs, and UN agencies. 6 49 7,330 1,446 6 3 deaths injured displaced people Affected houses Affected schools Health centers affected Situation Overview Tropical Cyclone Haruna made landfall over Morombe District on the south-west coast of Madagascar at 05:00 local time (UTC+3) on 22 February as a Category 2 tropical cyclone, with wind speeds of 154 km/h to 177 km/h. -

Madagascar: Tropical Cyclone Haruna Office of the Resident Coordinator Situation Report No
Madagascar: Tropical Cyclone Haruna Office of the Resident Coordinator Situation Report No. 2 (as of 23 Feb 2013) This report is produced by Office of the Resident Coordinator in Madagascar in collaboration with humanitarian partners. It covers the period from 22 to 23 February 2013. The next report will be issued around 25 February 2013. Highlights Tropical Cyclone Haruna made landfall over the south-west coast of Madagascar on 22 February 2013 as a powerful Category 2 Tropical Cyclone. Haruna made its exit to the Indian Ocean, as a tropical storm, around Taolagnaro at about 09:00 on 23 February. The most affected districts are Morombe and Toliary. Sakaraha and Betioky Sud are moderately affected. On 23 February, Fiherenana dyke burst, causing flooding to a large area and 25,000 people potentially needed to be evacuated. To date, some 1,000 people have been evacuated to temporary sites and 5,000 others are being hosted by their relatives. Initial situation assessments by local authorities and humanitarian partners partially started on 23 February. Reports of significant damage to houses, public buildings and infrastructure are being received. Aerial assessments will be conducted on 24 and 25 February by the Government, INGOs, and UN agencies. 6 49 7,330 1,446 6 3 deaths injured displaced people Affected houses Affected schools Health centers affected Situation Overview Tropical Cyclone Haruna made landfall over Morombe District on the south-west coast of Madagascar at 05:00 local time (UTC+3) on 22 February as a Category 2 tropical cyclone, with wind speeds of 154 km/h to 177 km/h. -

Cyclone Haruna Makes Landfall in Madagascar 22 February 2013
Cyclone Haruna makes landfall in Madagascar 22 February 2013 NASA's Aqua satellite captured a visible image of Tropical Storm Haruna on Feb. 22 at 1105 UTC (6:05 a.m. EST) after it moved inland and its eye was completed cloud-filled. MODIS imagery revealed that the size of the eye was smaller since the storm made landfall. As a result of its interaction with land, Haruna's maximum sustained winds dropped to 80 knots by 1200 UTC (7 a.m. EST) on Feb. 22. Haruna's cloud- filled eye was centered near 23.3 south latitude and 44.2 east longitude, about 300 nautical miles southwest of Antananarivo, the capital and largest city in Madagascar. Haruna was moving to the south-southeast at 7 knots and dropping heavy rainfall along its path. Radar at 10:30 a.m. EST (15:30 UTC) showed that Haruna's rainfall extends from the town of Belo Tsiribihina on Madagascar's west-central coast, east to the city of Fianarantsoa, and to the town of Taolagnaro on the southeastern coast. The MODIS instrument aboard NASA's Aqua satellite captured this visible image of Tropical Storm Haruna on Feb. 22 at 1105 UTC (6:05 a.m. EST). Credit: NASA Goddard MODIS Rapid Response Team NASA's Aqua satellite passed over Cyclone Haruna after it made landfall in southwestern Madagascar. Haruna's center made landfall near Manombo, Madagascar around 0600 UTC (1 a.m. EST/U.S.) The METEO-7 satellite captured a visible image of Haruna at the time of landfall and showed that its This night-time image revealed Cyclone Haruna's eye had already become cloud-filled. -
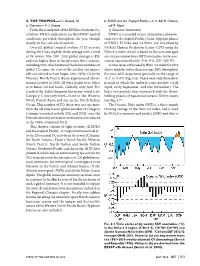
S133 2) Atmospheric Circulation on the Whole, Near-Average Atmo- Spheric Wind and Convection Patterns Reflected ENSO-Neutral Throughout the Year (Figs
4. THE TROPICS—H. J. Diamond, Ed. b. ENSO and the Tropical Pacific—G. D. Bell, M. L’Heureux, a. Overview—H. J. Diamond and M. Halpert From the standpoint of the El Niño-Southern Os- 1) OCEANIC CONDITIONS cillation (ENSO), indications are that ENSO-neutral ENSO is a coupled ocean-atmosphere phenom- conditions prevailed throughout the year, though enon over the tropical Pacific Ocean. Opposite phases mostly on the cool side of neutral. of ENSO, El Niño and La Niña, are classified by Overall, global tropical cyclone (TC) activity NOAA’s Climate Prediction Center (CPC) using the during 2013 was slightly above average with a total Niño-3.4 index, which is based on the area-averaged of 94 storms (the 1981–2010 global average is 89), sea surface temperature (SST) anomalies in the east- and was higher than in the previous three seasons, central equatorial Pacific (5°N–5°S, 170°–120°W). including 2010, which featured the lowest numbers of A time series of the weekly Niño-3.4 index for 2013 global TCs since the start of the satellite era (gener- shows slightly cooler-than-average SSTs throughout ally considered to have begun after 1970). Only the the year, with departures generally in the range of Western North Pacific Basin experienced above- -0.2° to -0.4°C (Fig. 4.1a). There were only three short normal activity in 2013; all other basins were either periods in which the index became positive (early at or below normal levels. Globally, only four TCs April, early September, and late November). -

Tropical Cyclone HARUNA in the Area of Madagascar in February 2013 Dr
Tropical Cyclone HARUNA in the area of Madagascar in February 2013 Dr. Susanne Haeseler; updated: 26 February 2013 Introduction and storm track On 20 February 2013, the tropical cyclone HARUNA had formed across the Mozambique Channel, the sea area between Africa and Madagascar (Fig. 1, 2 and 3a). On its way to the southern part of Madagascar's west coast, maximum sustained wind speeds (1 minute aver- age) up to about 100 knots (185 km/h) were reached. This is equivalent to a category 3 hur- ricane on the 1 to 5 rating Saffir-Simpson hurricane scale. On 22 February at about 02:30 UTC, the tropical cyclone made landfall about 55 km south of Morombe and crossed southern Madagascar while weakening (Fig. 3b). Heavy precipitation triggered by HARUNA caused regionally serious floods, and high wind speeds contributed to severe damage. Media reported of at least 10 deaths. In the early morning of 23 February, after having crossed the island in about 24 hours, the storm centre reached the eastern coast of Madagascar (Fig. 3c). East of the island, HARUNA re-strengthened to a tropical storm (tempête tropicale) for a short time, and then dissipated by and by. Fig. 1: Map of Madagascar and surroundings with stations mentioned below. 1 Fig. 2: Track of tropical cyclone HARUNA. Observation (solid line) and forecast (dashed line) on 24 February 2013. [Source: Meteo France; supplemented] 2 Wind and precipitation On Ile Europa, an island in the Mozambique Channel being affected by HARUNA, a mean wind speed (10 minute average) of 83 km/h (45 kn) was recorded on 20 February 2013, 12 UTC. -

Building Urban Resilience to Climate Change a Review of Madagascar
TECHNICAL REPORT BUILDING URBAN RESILIENCE TO CLIMATE CHANGE A REVIEW OF MADAGASCAR March 2018 This document was produced for review by the United States Agency for International Development. It was prepared by Chemonics for the Climate Change Adaptation, Thought Leadership and Assessments (ATLAS) Task Order No. AID- OAA-I-14-00013, under the Restoring the Environment through Prosperity, Livelihoods, and Conserving Ecosystems (REPLACE) IDIQ. Chemonics contact: Chris Perine, Chief of Party ([email protected]) Chemonics International Inc. 1717 H Street NW Washington, DC 20006 ATLAS reports and other products are available on the Climatelinks website: https://www.climatelinks.org/projects/atlas Cover Photo: Maria Olsen, European Union/European Commission Humanitarian and Civil Protection (EU/ECHO). The local Red Cross assists communities temporarily displaced by floods in Antananarivo’s Ankasina neighborhood. In 2015, Tropical Cyclone Chedza caused Antananarivo’s primary pumping station to cease operation, leading to flooding and contamination of water supply and sanitation infrastructures across Madagascar’s capital. BUILDING URBAN RESILIENCE TO CLIMATE CHANGE A REVIEW OF MADAGASCAR March 2018 Prepared for: United States Agency for International Development Climate Change Adaptation, Thought Leadership and Assessments (ATLAS) Prepared by: Veronique Lee and Fernanda Zermoglio (Chemonics International Inc.) This report is made possible by the support of the American people through the United States Agency for International Development