Why Are Plants Carnivorous?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

"National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary."
Intro 1996 National List of Vascular Plant Species That Occur in Wetlands The Fish and Wildlife Service has prepared a National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary (1996 National List). The 1996 National List is a draft revision of the National List of Plant Species That Occur in Wetlands: 1988 National Summary (Reed 1988) (1988 National List). The 1996 National List is provided to encourage additional public review and comments on the draft regional wetland indicator assignments. The 1996 National List reflects a significant amount of new information that has become available since 1988 on the wetland affinity of vascular plants. This new information has resulted from the extensive use of the 1988 National List in the field by individuals involved in wetland and other resource inventories, wetland identification and delineation, and wetland research. Interim Regional Interagency Review Panel (Regional Panel) changes in indicator status as well as additions and deletions to the 1988 National List were documented in Regional supplements. The National List was originally developed as an appendix to the Classification of Wetlands and Deepwater Habitats of the United States (Cowardin et al.1979) to aid in the consistent application of this classification system for wetlands in the field.. The 1996 National List also was developed to aid in determining the presence of hydrophytic vegetation in the Clean Water Act Section 404 wetland regulatory program and in the implementation of the swampbuster provisions of the Food Security Act. While not required by law or regulation, the Fish and Wildlife Service is making the 1996 National List available for review and comment. -

Quite a Few Reasons for Calling Carnivores 'The Most Wonderful
Annals of Botany 109: 47–64, 2012 doi:10.1093/aob/mcr249, available online at www.aob.oxfordjournals.org REVIEW Quite a few reasons for calling carnivores ‘the most wonderful plants in the world’ Elz˙bieta Kro´l1,*,†, Bartosz J. Płachno2,†, Lubomı´r Adamec3, Maria Stolarz1, Halina Dziubin´ska1 and Kazimierz Tre˛bacz1 1Department of Biophysics, Institute of Biology, Maria Curie-Skłodowska University, Akademicka 19, 20-033 Lublin, Poland, 2Department of Plant Cytology and Embryology, Jagiellonian University, Grodzka 52, 31-044 Cracow, Poland and 3Institute of Botany AS CR, Dukelska´ 135, 37982 Trˇebonˇ, Czech Republic †These authors contributed equally to this work. * For correspondence. E-mail [email protected] Received: 30 May 2011 Returned for revision: 28 June 2011 Accepted: 8 August 2011 Published electronically: 21 September 2011 Downloaded from † Background A plant is considered carnivorous if it receives any noticeable benefit from catching small animals. The morphological and physiological adaptations to carnivorous existence is most complex in plants, thanks to which carnivorous plants have been cited by Darwin as ‘the most wonderful plants in the world’. When considering the range of these adaptations, one realizes that the carnivory is a result of a multitude of different features. † Scope This review discusses a selection of relevant articles, culled from a wide array of research topics on plant carnivory, and focuses in particular on physiological processes associated with active trapping and digestion of http://aob.oxfordjournals.org/ prey. Carnivory offers the plants special advantages in habitats where nutrient supply is scarce. Counterbalancing costs are the investments in synthesis and the maintenance of trapping organs and hydrolysing enzymes. -

The First Record of the Boreal Bog Species Drosera Rotundifolia (Droseraceae) from the Philippines, and a Key to the Philippine Sundews
Blumea 61, 2016: 24–28 www.ingentaconnect.com/content/nhn/blumea RESEARCH ARTICLE http://dx.doi.org/10.3767/000651916X691330 The first record of the boreal bog species Drosera rotundifolia (Droseraceae) from the Philippines, and a key to the Philippine sundews F.P. Coritico1, A. Fleischmann2 Key words Abstract Drosera rotundifolia, a species of the temperate Northern Hemisphere with a disjunct occurrence in high montane West Papua, has been discovered in a highland peat bog on Mt Limbawon, Pantaron Range, Bukidnon carnivorous plants on the island of Mindanao, Philippines, which mediates to the only other known tropical, Southern Hemisphere Drosera location in New Guinea and the closest known northern populations in southern Japan and south-eastern China. Droseraceae A dichotomous key to the seven Drosera species of the Philippines is given, and distribution maps are provided. Malesia Mindanao Published on 15 March 2016 Northern Hemisphere - Tropics disjunction Philippines INTRODUCTION Drosera rotundifolia L. (the generic type) is a temperate, winter dormant species that is widespread in the Northern The Philippines are rich in carnivorous plants, with about 47 Hemisphere, from Pacific North America across large parts of species known from the islands, most of which belong to the northern America and Europe to Siberia and the Kamchatka pitcher plant genus Nepenthes L. This genus has more than 30 Peninsula, South Korea and Japan. It is the Drosera spe- species in the Philippines, all except Nepenthes mirabilis (Lour.) cies covering the largest range, spanning the entire Northern Druce endemic to the country. Most species occur on Mindanao Hemisphere from 180° Western Longitude to about 180° East, and Palawan, while several are confined to a single highland however, not forming a continuous circumboreal range (Diels or even mountain peak (Robinson et al. -

Carnivorous Plant Responses to Resource Availability
Carnivorous plant responses to resource availability: environmental interactions, morphology and biochemistry Christopher R. Hatcher A doctoral thesis submitted in partial fulfilment of requirements for the award of Doctor of Philosophy of Loughborough University November 2019 © by Christopher R. Hatcher (2019) Abstract Understanding how organisms respond to resources available in the environment is a fundamental goal of ecology. Resource availability controls ecological processes at all levels of organisation, from molecular characteristics of individuals to community and biosphere. Climate change and other anthropogenically driven factors are altering environmental resource availability, and likely affects ecology at all levels of organisation. It is critical, therefore, to understand the ecological impact of environmental variation at a range of spatial and temporal scales. Consequently, I bring physiological, ecological, biochemical and evolutionary research together to determine how plants respond to resource availability. In this thesis I have measured the effects of resource availability on phenotypic plasticity, intraspecific trait variation and metabolic responses of carnivorous sundew plants. Carnivorous plants are interesting model systems for a range of evolutionary and ecological questions because of their specific adaptations to attaining nutrients. They can, therefore, provide interesting perspectives on existing questions, in this case trait-environment interactions, plant strategies and plant responses to predicted future environmental scenarios. In a manipulative experiment, I measured the phenotypic plasticity of naturally shaded Drosera rotundifolia in response to disturbance mediated changes in light availability over successive growing seasons. Following selective disturbance, D. rotundifolia became more carnivorous by increasing the number of trichomes and trichome density. These plants derived more N from prey and flowered earlier. -

Carnivorous Plant Newsletter V42 N3 September 2013
Technical Refereed Contribution Phylogeny and biogeography of the Sarraceniaceae JOHN BRITTNACHER • Ashland, Oregon • USA • [email protected] Keywords: History: Sarraceniaceae evolution The carnivorous plant family Sarraceniaceae in the order Ericales consists of three genera: Dar- lingtonia, Heliamphora, and Sarracenia. Darlingtonia is represented by one species that is found in northern California and western Oregon. The genus Heliamphora currently has 23 recognized species all of which are native to the Guiana Highlands primarily in Venezuela with some spillover across the borders into Brazil and Guyana. Sarracenia has 15 species and subspecies, all but one of which are located in the southeastern USA. The range of Sarracenia purpurea extends into the northern USA and Canada. Closely related families in the plant order Ericales include the Roridu- laceae consisting of two sticky-leaved carnivorous plant species, Actinidiaceae, the Chinese goose- berry family, Cyrillaceae, which includes the common wetland plant Cyrilla racemiflora, and the family Clethraceae, which also has wetland plants including Clethra alnifolia. The rather charismatic plants of the Sarraceniaceae have drawn attention since the mid 19th century from botanists trying to understand how they came into being, how the genera are related to each other, and how they came to have such disjunct distributions. Before the advent of DNA sequencing it was very difficult to determine their relationships. Macfarlane (1889, 1893) proposed a phylogeny of the Sarraceniaceae based on his judgment of the overlap in features of the adult pitchers and his assumption that Nepenthes is a member of the family (Fig. 1a). He based his phy- logeny on the idea that the pitchers are produced from the fusion of two to five leaflets. -

Universidade Estadual Paulista Câmpus De
Campus de Botucatu UNESP - UNIVERSIDADE ESTADUAL PAULISTA CÂMPUS DE BOTUCATU INSTITUTO DE BIOCIÊN CIAS GENÔMICA ORGANELAR E EVOLUÇÃO DE GENLISEA E UTRICULARIA (LENTIBULARIACEAE) SAURA RODRIGUES DA SILVA Tese apresentada ao Instituto de Biociências, Câmpus de Botucatu, UNESP, para obtenção do título de Doutor em Ciências Biológicas (Botânica) BOTUCATU - SP - 2018 - Insti tuto de Biociências – Departamento de Botânica Distrito de Rubião Júnior s/n CEP 18618 - 000 Botucatu SP Brasil Tel 14 3811 6265/6053 fax 14 3815 3744 [email protected] Campus de Botucatu UNESP - UNIVERSIDADE ESTADUAL PAULISTA CÂMPUS DE BOTUCATU INSTITUTO DE BIOCIÊN CIAS GENÔMICA ORGANELAR E EVOLUÇÃO DE GENLISEA E UTRICULARIA (LENTIBULARIACEAE) SAUR A RODRIGUES DA SILVA PROF. DR. VITOR FERN ANDES OLIVEIRA DE MI RANDA ORIENTADOR PROF. DR. ALESSANDRO DE MEL L O VARANI Coorientador Tese apresentada ao Instituto de Biociências, Câmpus de Botucatu, UNESP, para obtenção do título de Doutor em Ciên cias Biológicas (Botânica) BOTUCATU - SP - 2018 - Instituto de Biociências – Departamento de Botânica Distrito de Rubião Júnior s/n CEP 18618 - 000 Botucatu SP Brasil Tel 14 3811 6265/6053 fax 14 3815 3744 [email protected] 2 Campus de Botucatu FICHA CATALOGRÁFIC A ELABORADA PELA SEÇÃO TÉCNICA DE AQUISIÇÃO E TRATAMENTO DA INFORMAÇÃO DIVISÃO TÉCNICA DE BIBLIOTECA E DOCUMENTAÇÃO - CAMPUS DE BOTUCATU - UNESP Silva, Saura Rodrigues. Genômica organelar e evolução d e Genlisea e Utricularia (Lentibulariaceae) / Saura Rodrigues da Silva. – 2018. Tese (doutorado) – Universidade Estadual Paulista, Instituto de Biociências de Botucatu, 2018. Orientador: Vitor Fernandes Oliveira de Miranda Co - orientador: Alessandro de Mello Varani Assunto CAPES: 1. Sistemática Vegetal CDD 581.1 Palavras - c have: Utricularia ; Genlisea ; genômica de organelas; ndhs; Evolução de organelas. -

Florida Council of Bromeliad Societies, Inc
Florida Council of Bromeliad Societies, Inc. In This Issue: 2007 Shows and Sales Cold Hardy Bromeliads List Vol. 27 Issue 1 February 2007 FCBS Affiliated Societies and Representatives B. Guild Tampa Bay Caloosahatchee Tom Wolfe Vicky Chirnside 5211 Lake LeClare Road 951 Southland Road Lutz 33558 Venice 34293 813-961-1475 941-493-5825 [email protected] [email protected] Bob Teems Tom Foley 813-855-0938 239-458-4656 Broward County Fl. East Coast Jose Donayre Calandra Thurrott 1240 Jefferson St. 713 Breckenridge Drive Hollywood 33019-1807 Port Orange 32127 954-925-5112 386-761-4804 Jcadonayre @bellsouth.net [email protected] Colleen Hendrix Carolyn Schoenau 954-530-0076 352-372-6589 Central Florida F. West Coast Betsy McCrory Linda Sheetz 3615 Boggy Creek Rd. 1153 Williams Dr. S Kissimmee 34744 St. Petersburg 33705 407-348-2139 727-864-3165 [email protected] [email protected] Butch Force Brian Corey 407-886-4814 727-864-3165 South Florida Gainesville Juan Espinosa-Almodovar Al Muzzell P.O. Box 430722 P.O. Box 14442 Miami 33243 Gainesville 32604 305-667-6155 352-372-4576 [email protected] John R. Moxley Michael Michalski 352-528-0783 305-279-2416 (Continued on the inside back cover.) 2007 Bromeliad Extravaganza Presented by Florida Council of Bromeliad Societies Hosted by the Bromeliad Society of Broward County Saturday, September 29, 2007 at the Hilton Ft. Lauderdale Airport Hotel 1870 Griffin Rd. Dania Beach, FL 33004 954-920-3300 954-920-3348 (fax) Room rates: Single or double $89.00 Rates in effect until September 14, 2007 Sale, Banquet, Raffle and Rare Plant Auction will take place at the same location. -

GREAT PLAINS REGION - NWPL 2016 FINAL RATINGS User Notes: 1) Plant Species Not Listed Are Considered UPL for Wetland Delineation Purposes
GREAT PLAINS REGION - NWPL 2016 FINAL RATINGS User Notes: 1) Plant species not listed are considered UPL for wetland delineation purposes. 2) A few UPL species are listed because they are rated FACU or wetter in at least one Corps region. -

A New Carnivorous Plant Lineage (Triantha) with a Unique Sticky-Inflorescence Trap
A new carnivorous plant lineage (Triantha) with a unique sticky-inflorescence trap Qianshi Lina,b,1, Cécile Anéc,d, Thomas J. Givnishc, and Sean W. Grahama,b aDepartment of Botany, University of British Columbia, Vancouver, BC V6T 1Z4, Canada; bUBC Botanical Garden, University of British Columbia, Vancouver, BC V6T 1Z4, Canada; cDepartment of Botany, University of Wisconsin–Madison, Madison, WI 53706; and dDepartment of Statistics, University of Wisconsin–Madison, Madison WI 53706 Edited by Elizabeth A. Kellogg, Donald Danforth Plant Science Center, St. Louis, MO, and approved June 5, 2021 (received for review October 30, 2020) Carnivorous plants consume animals for mineral nutrients that and in wetlands, including bogs, marly shorelines, and calcareous enhance growth and reproduction in nutrient-poor environments. spring-fed fens. In bogs, T. occidentalis is commonly found with Here, we report that Triantha occidentalis (Tofieldiaceae) represents recognized carnivorous plants such as Drosera rotundifolia a previously overlooked carnivorous lineage that captures insects on (Droseraceae) and Pinguicula vulgaris (Lentibulariaceae). During sticky inflorescences. Field experiments, isotopic data, and mixing the summer flowering season, T. occidentalis produces leafless models demonstrate significant N transfer from prey to Triantha, erect flowering stems up to 80 cm tall (12). These scapes have with an estimated 64% of leaf N obtained from prey capture in sticky glandular hairs, especially on their upper portions, a feature previous years, comparable to levels inferred for the cooccurring distinguishing Triantha from other genera of Tofieldiaceae round-leaved sundew, a recognized carnivore. N obtained via carnivory (Fig. 1). Small insects are often found trapped by these hairs; is exported from the inflorescence and developing fruits and may herbarium specimens are frequently covered in insects (Fig. -

National List of Vascular Plant Species That Occur in Wetlands 1996
National List of Vascular Plant Species that Occur in Wetlands: 1996 National Summary Indicator by Region and Subregion Scientific Name/ North North Central South Inter- National Subregion Northeast Southeast Central Plains Plains Plains Southwest mountain Northwest California Alaska Caribbean Hawaii Indicator Range Abies amabilis (Dougl. ex Loud.) Dougl. ex Forbes FACU FACU UPL UPL,FACU Abies balsamea (L.) P. Mill. FAC FACW FAC,FACW Abies concolor (Gord. & Glend.) Lindl. ex Hildebr. NI NI NI NI NI UPL UPL Abies fraseri (Pursh) Poir. FACU FACU FACU Abies grandis (Dougl. ex D. Don) Lindl. FACU-* NI FACU-* Abies lasiocarpa (Hook.) Nutt. NI NI FACU+ FACU- FACU FAC UPL UPL,FAC Abies magnifica A. Murr. NI UPL NI FACU UPL,FACU Abildgaardia ovata (Burm. f.) Kral FACW+ FAC+ FAC+,FACW+ Abutilon theophrasti Medik. UPL FACU- FACU- UPL UPL UPL UPL UPL NI NI UPL,FACU- Acacia choriophylla Benth. FAC* FAC* Acacia farnesiana (L.) Willd. FACU NI NI* NI NI FACU Acacia greggii Gray UPL UPL FACU FACU UPL,FACU Acacia macracantha Humb. & Bonpl. ex Willd. NI FAC FAC Acacia minuta ssp. minuta (M.E. Jones) Beauchamp FACU FACU Acaena exigua Gray OBL OBL Acalypha bisetosa Bertol. ex Spreng. FACW FACW Acalypha virginica L. FACU- FACU- FAC- FACU- FACU- FACU* FACU-,FAC- Acalypha virginica var. rhomboidea (Raf.) Cooperrider FACU- FAC- FACU FACU- FACU- FACU* FACU-,FAC- Acanthocereus tetragonus (L.) Humm. FAC* NI NI FAC* Acanthomintha ilicifolia (Gray) Gray FAC* FAC* Acanthus ebracteatus Vahl OBL OBL Acer circinatum Pursh FAC- FAC NI FAC-,FAC Acer glabrum Torr. FAC FAC FAC FACU FACU* FAC FACU FACU*,FAC Acer grandidentatum Nutt. -
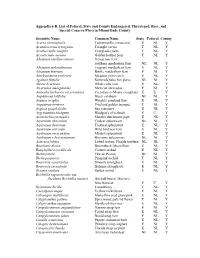
Conservation Appendix 6-B Listed Flora
Appendix 6-B. List of Federal, State and County Endangered, Threatened, Rare, and Special Concern Flora in Miami-Dade County Scientific Name Common Name State Federal County Acacia choriophylla Tamarindillo; cinnecord E NL Y Acanthocereus tetragenus Triangle cactus T NL Y Acoelorraphe wrightii Everglades palm T NL Y Acrostichum aureum Golden leather fern T NL Y Adiantum capillus-veneris Venus hair fern; southern maidenhair fern NL NL Y Adiantum melanoleucum Fragrant maidenhair fern E NL Y Adiantum tenerum Brittle maidenhair fern E NL Y Aeschynomene pratensis Meadow joint-vetch E NL Y Agalinis filifolia Seminole false fox glove NL NL Y Aletris bracteata White colic root E NL Y Alvaradoa amorphoides Mexican alvaradoa E NL Y Amorpha herbacea var.crenulata Crenulate (=Miami) leadplant E E Y Amphitecna latifolia Black calabash NL NL Y Anemia wrightii Wright's pineland fern E NL Y Angadenia berteroi Pineland golden trumpet T NL Y Argusia gnaphalodes Sea rosemary E NL Y Argythamnia blodgettii Blodgett's silverbush E C Y Aristolochia pentandra Marsh's dutchmans pipe E NL Y Asplenium abscissum Cutleaf spleenwort NL NL Y Asplenium dentatum Toothed spleenwort E NL Y Asplenium serratum Wild bird nest fern E NL Y Asplenium verecundum Modest spleenwort E NL Y Asplenium x biscaynianum Biscayne spleenwort NL NL Y Asteraea lobata Lobed croton; Florida treefern NL NL Y Baccharis dioica Broombush falsewillow E NL Y Basiphyllaea corallicola Carter's orchid E NL Y Bletia patula Flor de Pesmo NL NL Y Bletia purpurea Pinepink orchid T NL Y Bourreria cassinifolia Smooth strongback E NL Y Bourreria succulenta Bahama strongback E NL Y Brassia caudata Spider orchid E NL Y Brickellia eupatorioides var. -

Mountain Plants of Northeastern Utah
MOUNTAIN PLANTS OF NORTHEASTERN UTAH Original booklet and drawings by Berniece A. Andersen and Arthur H. Holmgren Revised May 1996 HG 506 FOREWORD In the original printing, the purpose of this manual was to serve as a guide for students, amateur botanists and anyone interested in the wildflowers of a rather limited geographic area. The intent was to depict and describe over 400 common, conspicuous or beautiful species. In this revision we have tried to maintain the intent and integrity of the original. Scientific names have been updated in accordance with changes in taxonomic thought since the time of the first printing. Some changes have been incorporated in order to make the manual more user-friendly for the beginner. The species are now organized primarily by floral color. We hope that these changes serve to enhance the enjoyment and usefulness of this long-popular manual. We would also like to thank Larry A. Rupp, Extension Horticulture Specialist, for critical review of the draft and for the cover photo. Linda Allen, Assistant Curator, Intermountain Herbarium Donna H. Falkenborg, Extension Editor Utah State University Extension is an affirmative action/equal employment opportunity employer and educational organization. We offer our programs to persons regardless of race, color, national origin, sex, religion, age or disability. Issued in furtherance of Cooperative Extension work, Acts of May 8 and June 30, 1914, in cooperation with the U.S. Department of Agriculture, Robert L. Gilliland, Vice-President and Director, Cooperative Extension