03 Chapters 3 Evans
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
Additional Information
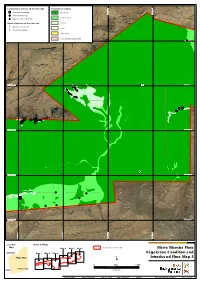
Current Survey Introduced Flora Records Vegetation Condition *Acetosa vesicaria Excellent 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 mE 535,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 536,000 mE 536,000 537,000 mE 537,000 534,000 mE 534,000 mE 535,000 534,000 mE 534,000 -

Wiregrass (Aristida Stricta)
Wiregrass (Aristida stricta) For definitions of botanical terms, visit en.wikipedia.org/wiki/Glossary_of_botanical_terms. Wiregrass is a perennial bunchgrass found in scrub, pinelands and coastal uplands throughout much of Florida. It is the dominant groundcover species in longleaf pine savannas and is a primary food source for gopher tortoises. Birds and small wildlife eat the seeds. Historically, cattle grazed on Wiregrass’s tender new growth. Wiregrass flowers are tiny and brown. They are born on spikelike terminal panicles Flower stalks are elongated and extend above the leaves. Leaf blades are long, thin and rolled inward, giving them a wiry appearance (hence the common name). They are erect and green when young Photo courtesy of Alan Cressler, Lady Bird Johnson Wildflower Center and begin to arch and turn brown as they age. Its fruits are small, yellowish caryopses. Seeds may be dispersed by wind, gravity or on the fur of passing animals. The genus name Aristida is from the Latin arista, meaning “awn” and referring to the three awns or bristle-like structures that extend from the florets. (An alternative common name is Pineland threeawn.) The species epithet, stricta, is from the Latin strictus, meaning straight or erect. Family: Poaceae (Grass family) Native range: Nearly throughout To see where natural populations of Wiregrass have been vouchered, visit www.florida.plantatlas.usf.edu. Hardiness: Zones 8A–10B Lifespan: Perennial Soil: Moist to dry, well-drained sandy soils Exposure: Full sun to partial shade Growth habit: 1–3’+ tall and equally wide Propagation: Division, seed Garden tips: Wiregrass is fast-growing and tolerant of drought conditions and low-nutrient soils. -

Wildlife Preservation Winter 2011.Indd
Linda Dennis, Regional Advisor WPSA The northern hairy-nosed wombat (Lasiorhinus krefftii) is one of Australia’s most mysterious animals. Only a hundred or so people have been lucky enough to see one in the wild, and there are none in captivity anywhere in the world. Predator proof fence around Epping Forest. Photo: Linda Dennis In recent years, however, the northern it makes it into the Top 10 of the Xstrata donated a massive $3 million hairy-nosed wombat has become world’s most endangered animals. to assist in the translocation of just that little bit more known in some wombats from Epping Forest Australia and around the world – Every two or so years a northern to the Richard Underwood Nature but unfortunately for all the wrong hairy-nosed hair census is carried out. Refuge, near St George in southern reasons. It is Australia’s second most Volunteers from all corners of Australia Queensland to form a second colony. endangered animal, the rst being – and even the world – converge on the Gilbert’s potoroo of Western Epping Forest National Park (Scienti c) A brief history of the northern Australia. in central Queensland for two weeks to hairy-nosed wombat walk the length of the park, collecting The northern hairy-nosed wombat wombat hair, measuring footprints and Fossil records show that the has the dubious honour of making it counting wombat poo. northern hairy-nosed wombat was to more than one endangered listing: once widespread, living in Victoria, The results of the 2007 hair census New South Wales and Queensland • Endangered in Queensland proclaimed that there are only 138 (DERM). -

Department of the Interior Fish and Wildlife Service
Tuesday, February 10, 2009 Part II Department of the Interior Fish and Wildlife Service 50 CFR Part 17 Endangered and Threatened Wildlife and Plants; Determination of Endangered Status for Reticulated Flatwoods Salamander; Designation of Critical Habitat for Frosted Flatwoods Salamander and Reticulated Flatwoods Salamander; Final Rule VerDate Nov<24>2008 14:17 Feb 09, 2009 Jkt 217001 PO 00000 Frm 00001 Fmt 4717 Sfmt 4717 E:\FR\FM\10FER2.SGM 10FER2 erowe on PROD1PC63 with RULES_2 6700 Federal Register / Vol. 74, No. 26 / Tuesday, February 10, 2009 / Rules and Regulations DEPARTMENT OF THE INTERIOR during normal business hours, at U.S. Register on or before July 30, 2008, with Fish and Wildlife Service, Mississippi the final critical habitat rule to be Fish and Wildlife Service Fish and Wildlife Office, 6578 Dogwood submitted for publication in the Federal View Parkway, Jackson, MS 39213. Register by January 30, 2009. The 50 CFR Part 17 FOR FURTHER INFORMATION CONTACT: Ray revised proposed rule was signed on [FWS–R4–ES–2008–0082; MO 9921050083– Aycock, Field Supervisor, U.S. Fish and and delivered to the Federal Register on B2] Wildlife Service, Mississippi Field July 30, 2008, and it subsequently Office, 6578 Dogwood View Parkway, published on August 13, 2008 (73 FR RIN 1018–AU85 Jackson, MS 39213; telephone: 601– 47258). We also published supplemental information on the Endangered and Threatened Wildlife 321–1122; facsimile: 601–965–4340. If proposed rule to maintain the status of and Plants; Determination of you use a telecommunications device the frosted flatwoods salamander as Endangered Status for Reticulated for the deaf (TDD), call the Federal Information Relay Service (FIRS) at threatened (73 FR 54125; September 18, Flatwoods Salamander; Designation of 2008). -

Jan Dvorak (As Quickly Written Down by a Person with Poor Hearing…Me)
1/22/2018 The Impact of Polyploidy on Genome Evolution in Poales and Other Monocots “I don’t have to emphasize that gene duplications are the fabric of evolution in plants.” -Jan Dvorak (as quickly written down by a person with poor hearing…me) Michael R. McKain The University of Alabama @mrmckain @mrmckain Poales Diversity Grass genomes: the choose your own adventure of genome evolution • ~22,800 species • ~11,088 species in Poaceae • Transposons (McClintock, Wessler) • GC content bias (Carels and Bernardi 2000) • Three WGD events 0 0 4 0 0 • rho (Peterson et al. 2004) 3 y c n 0 e 0 u 2 q e r F • 0 sigma (Tang et al. 2010) 0 1 • tau (Tang et al. 2010, Jiao et al. 2014) 0 %GC Givnish et al. 2010 @mrmckain Schnable et al., 2009 Zeroing in on WGD placement Banana genome Pineapple genome How has ancient polyploidy altered the genomic landscape in grasses and other Poales? D’Hont et al. 2012 Ming, VanBuren et al. 2015 Recovered sigma after grass divergence from commelinids Recovered sigma after grass+pineapple divergence from commelinids @mrmckain @mrmckain 1 1/22/2018 Phylotranscriptomic approach Coalescence-based Phylogeny of 234 Single-copy genes • Sampling 27 transcriptomes and 7 genomes • Phylogeny consistent with previous • Representation for all families (except Thurniaceae) in nuclear gene results Poales • Conflicting topology with • RNA from young leaf or apical meristem, a combination of chloroplast genome tree: Moncot Tree of Life and 1KP • Ecdeiocolea/Joinvillea sister instead of • General steps: a grade • Trinity assembly • Typha -

List of Plant Species List of Plant Species
List of plant species List of Plant Species Contents Amendment history .......................................................................................................................... 2 1 Introduction ...................................................................................................................................... 3 1.1 Application ........................................................................................................................... 3 1.2 Relationship with planning scheme ..................................................................................... 3 1.3 Purpose ............................................................................................................................... 3 1.4 Aim ...................................................................................................................................... 3 1.5 Who should use this manual? ............................................................................................. 3 2 Special consideration ....................................................................................................................... 3 3 Variations ......................................................................................................................................... 4 4 Relationship ..................................................................................................................................... 4 Appendix A – Explanatory notes & definitions ....................................................................................... -

Pasture Condition Guide for the Ord River Catchment
Bulletin 4769 Department of June 2009 Agriculture and Food ISSN 1833-7236 Pasture condition guide for the Ord River Catchment Department of Agriculture and Food Pasture condition guide for the Ord River Catchment K. Ryan, E. Tierney & P. Novelly Copyright © Western Australian Agriculture Authority, 2009 Acknowledgements Photographs by S. Eyres and the Department of Agriculture and Food, Western Australia (DAFWA) Photographic Unit The information in this publication has been developed in consultation with experienced rangelands field staff providing services to the East Kimberley pastoral leases and with reference to Range Condition Guides for the West Kimberley Area, WA (Payne, Kubicki and Wilcox 1974) and Lands of the Ord–Victoria Area, WA and NT (Stewart et al. 1970). The authors would like to thank all those who provided valuable feedback on the design and content of this guide, including Andrew Craig, David Hadden and Matthew Fletcher (DAFWA Kununurra), Simon Eyres (DAFWA Photographic Unit), Alan Payne (retired DAFWA rangelands advisor), and members of the Halls Creek—East Kimberley Land Conservation District. This project was funded by Rangelands NRM WA using National Action Plan for Salinity and Water Quality funding. Rangelands NRM WA regards this project as a strategic investment which will contribute to an improved understanding and awareness of pasture condition in the Ord Catchment, leading to improved land management in that area. Rangelands NRM WA contracted the Department of Agriculture and Food WA to undertake the project. Funding for the National Action Plan for Salinity and Water Quality was provided by the Australian and Western Australian Governments. Disclaimer The Chief Executive Officer of the Department of Agriculture and Food and the State of Western Australia accept no liability whatsoever by reason of negligence or otherwise arising from the use or release of this information or any part of it. -
The Nursery Industry - Protecting Our Environment 1) Contents
A Guide for Gardeners in South Queensland South West West The Nursery Industry - Protecting Our Environment 1) Contents 1. Contents 2. Area Map 3. Acknowledgements 4. Introduction from the Ministers 5. Introduction 6. What is Grow Me Instead? 7. Establishing the criteria for Grow Me Instead 8. What you can do! 9. Controlling weeds 10. The invasive plants and their alternatives Trees and Shrubs Climbing and Ground Cover Plants Berried Plants Grasses Bulbous Plants Succulent Plants Aquatic Plants 11 . Invasive plant index 12 . Further information and additional resources The publication of the Grow Me Instead Queensland South West guide has been generously supported by Queensland Murray ‐ Darling Committee and Tara and District Landcare. Funded by: 2 Cover photo courtesy of Queensland Murray - Darling Committee 2) Area Map The publication of the Grow Me Instead Queensland South West guide has been generously supported by South West NRM Ltd. 3 3) Acknowledgements The Grow Me Instead South West project has been completed by NGIQ through the generous funding support of the following organisations; Funded by: 4 The ‘Grow Me Instead’ (GMI) project was first initiated by the Nursery and Garden Industry of NSW & ACT (NGINA) as a voluntary partnership with its member nurseries to encourage the removal from production and sale of plants known to be invasive in the natural environment. The original Grow Me Instead Queensland project was supported by funding from Australian Government NRM Team GPO Box 787 Canberra ACT 2601 Australia Ph 1 800 552008 (Toll Free) Web www.nrm.gov.au Special thanks go to Grow Me Instead project coordinator Barry Naylor (NGIQ) and project manager John McDonald (Nursery Industry Development Manager NGIQ). -

Genome Comparison Reveals Mutation Hotspots in the Chloroplast Genome and Phylogenetic Relationships of Ormosia Species
Hindawi BioMed Research International Volume 2019, Article ID 7265030, 11 pages https://doi.org/10.1155/2019/7265030 Research Article Genome Comparison Reveals Mutation Hotspots in the Chloroplast Genome and Phylogenetic Relationships of Ormosia Species Hongshan Liu,1,2 Zhihai Su,2 Shuiqing Yu,2 Jialin Liu,2 Xiaojuan Yin,2 Guowei Zhang,2 Wei Liu,2 and Bin Li 1 State Key Laboratory of Tree Breeding and Forest Genetics, Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration, Research Institute of Forestry, Chinese Academy of Forestry, Beijing , China Administration Bureau of Hongyashan State Owned Forest Farm of Hebei Province, Yixian , China Correspondence should be addressed to Bin Li; [email protected] Received 23 March 2019; Revised 13 July 2019; Accepted 22 July 2019; Published 21 August 2019 Academic Editor: Gerald J. Wyckof Copyright © 2019 Hongshan Liu et al. Tis is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Te papilionoid legume genus Ormosia comprises approximately 130 species, which are distributed mostly in the Neotropics, with some species in eastern Asia and northeastern Australia. Te taxonomy and evolutionary history remain unclear due to the lack of a robust species-level phylogeny. Chloroplast genomes can provide important information for phylogenetic and population genetic studies. In this study, we determined the complete chloroplast genome sequences of fve Ormosia species by Illumina sequencing. Te Ormosia chloroplast genomes displayed the typical quadripartite structure of angiosperms, which consisted of a pair of inverted regions separated by a large single-copy region and a small single-copy region. -

Grasses of Namibia Contact
Checklist of grasses in Namibia Esmerialda S. Klaassen & Patricia Craven For any enquiries about the grasses of Namibia contact: National Botanical Research Institute Private Bag 13184 Windhoek Namibia Tel. (264) 61 202 2023 Fax: (264) 61 258153 E-mail: [email protected] Guidelines for using the checklist Cymbopogon excavatus (Hochst.) Stapf ex Burtt Davy N 9900720 Synonyms: Andropogon excavatus Hochst. 47 Common names: Breëblaarterpentyngras A; Broad-leaved turpentine grass E; Breitblättriges Pfeffergras G; dukwa, heng’ge, kamakama (-si) J Life form: perennial Abundance: uncommon to locally common Habitat: various Distribution: southern Africa Notes: said to smell of turpentine hence common name E2 Uses: used as a thatching grass E3 Cited specimen: Giess 3152 Reference: 37; 47 Botanical Name: The grasses are arranged in alphabetical or- Rukwangali R der according to the currently accepted botanical names. This Shishambyu Sh publication updates the list in Craven (1999). Silozi L Thimbukushu T Status: The following icons indicate the present known status of the grass in Namibia: Life form: This indicates if the plant is generally an annual or G Endemic—occurs only within the political boundaries of perennial and in certain cases whether the plant occurs in water Namibia. as a hydrophyte. = Near endemic—occurs in Namibia and immediate sur- rounding areas in neighbouring countries. Abundance: The frequency of occurrence according to her- N Endemic to southern Africa—occurs more widely within barium holdings of specimens at WIND and PRE is indicated political boundaries of southern Africa. here. 7 Naturalised—not indigenous, but growing naturally. < Cultivated. Habitat: The general environment in which the grasses are % Escapee—a grass that is not indigenous to Namibia and found, is indicated here according to Namibian records. -

Plants for Whitsunday Gardens
Whitsunday Plants [email protected] Suitable for Gardens Ph: (07) 4945 0267 What are Local Native Plants? Native plants are also far less Local native plants are those that are indigenous to the likely than exotic (non-native) Whitsunday region. There are many local native plants plants to become that can be used instead of exotic (introduced) plants or environmental weeds. Native even those native to other parts of Australia. Wedelia (pictured left) does not have the invasive qualities of the similar in appearance Why Use Local Native Plants? declared weed Singapore Local native plants are often better adapted to local Daisy. conditions and also provide a natural food source for Wedelia spilanthoides animals, birds and insects. It is important to plant a Photo: C. Peterson variety of plants to supply nectar, fruit and seeds to native animals. Things to Consider When Planning Below: Cajanus Your Garden: reticulatus, a low, Where space is limited such as in household gardens, dense shrub provides seeds to native under power lines and near driveways and buildings, animals. there is a need to use plants that will not become a Photo: P. Alden menace to neighbours or damage infrastructure. It is important to consider the mature size of plants, including width, height and how extensive the roots systems will become. Above: Melicope elleryana provides nectar, and later seeds to native Figs in particular (Ficus species) should not be planted animals. Photo: D. Pepplinkhouse near infrastructure. There are attractive grasses, ground covers, small to medium shrubs and small trees with colourful flowers, wonderful aromas or unusual fruit. -

Biological Opinion
Biological Opinion Francis Marion National Forest Revised Land Management Plan FWS Log No.04ES1000-2016-F-0628 U.S. Department of Agriculture Forest Service December 2, 2016 Prepared by: U.S. Fish and Wildlife Service South Carolina Ecological Services Field Office 176 Croghan Spur Road, Suite 200 Charleston, South Carolina 29407 LIST OF FIGURES FIGURE 1. Ecosystems (not including rivers and streams) on the Francis Marion National Forest. FIGURE 2. Management areas on the Francis Marion National Forest. FIGURE 3. Red-cockaded woodpecker (Picoides borealis) demographic populations on Francis Marion, spatially defined as aggregations of active clusters within 6 km (3.7 miles), and important areas (red arrows) to either establish or maintain demographic connectivity. FIGURE 4: Red-cockaded woodpecker (Picoides borealis) Francis Marion population size and trend, active clusters. Data and graph prepared Mark Danaher, FMNF, 2014. FIGURE 5: Red-cockaded woodpecker (Picoides borealis) Francis Marion population size and trend, potential breeding groups. Data and graph prepared Mark Danaher, FMNF, 2014. 2 LIST OF TABLES TABLE 1. Summary of Effects Determinations. TABLE 2. Fire-adapted ecosystems by Management Area. TABLE 3. Threatened or endangered species considered in this analysis. TABLE 4. American chaffseed (Schwalbea americana) monitoring trends from 1999-2016, Francis Marion National Forest. Data and table prepared by Robin Mackie, Forest Botanist/Ecologist, Francis Marion National Forest. TABLE 5. American chaffseed (Schwalbea americana) site prescribed burning trends, Francis Marion National Forest, 1996-2012. Data and table prepared by Robin Mackie, Forest Botanist/Ecologist, Francis Marion National Forest. TABLE 6. Roads and system type within Francis Marion managed cluster polygons.