Status of Coral Reefs in Trincomalee, Sri Lanka
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Part 5: List of Annexes
PART 5: LIST OF ANNEXES Annex 1: Letter of Endorsement Annex 2: Site Description and Maps Annex 3: Climate change Vulnerability and Adaptation Summary Annex 4: Incremental Cost Analysis Annex 5: Stakeholder Involvement Plan Annex 6: List of contacts Annex 7: Socioeconomic Status Report Annex 8: Monitoring and Evaluation Plan Annex 9: Bibliography Annex 10: Logical Framework Analysis Annex 11: Response to STAP Review Annex 12: Letter of Commitment- Coast Conservation Department Annex 13: Letter of Commitment- Ministry of Environment Annex 14: Letter of Commitment- International Fund for Agricultural Development _________________________________________________________________________________________________51 Tsunami Coastal Restoration in Eastern Sri Lanka Annex 2: Site Description and Maps Preamble The project is designed for the restoration and rehabilitation of coastal ecosystems. The initial emphasis of this five-year project will be on developing a scientifically based, low-cost, community-based approach to rehabilitating key coastal ecosystems at specific sites in the East Coast and facilitating replication of these techniques all along the East Coast (and in due course other tsunami-affected coasts). Three sites representing three major ecosystems – mangroves, coastal lagoons, and sand dunes –have been identified for piloting these themes. The selection was based on outputs from the Threats Analysis and the following criteria. 1. Hotspot analysis: sites where the tsunami effect was severe on the ecosystems and post tsunami reconstructions are in progress, global/national biodiversity importance exist, concentration of various resource users and their high dependency over the available resources exist and user conflicts exist. 2. Accessibility: accessibility by road was a criterion for selecting pilot sites 3. Absence of ongoing management and monitoring projects: sites at which on-going projects have not being considered for selection 4. -

Download Brochure
Redefining Beach Living The first of it’s kind, Oceanfront Condos presents a luxurious beachside option for those looking for a place to call home. Get away from bustling work life and enjoy gorgeous ocean views, spacious living spaces, and tranquil natural surroundings. With 54 condominiums in 4 identical building footprints and located only 260 kilometers away from the Capital of Sri Lanka, Oceanfront Condos is a home away from home. Regular Flights available from Colombo to Trincomalee - China Bay Sri Lanka Floor Plans 3 BEDROOM 2 BEDROOM TYPE A TYPE B 122.14 SQM 110.00 SQM 1314.23 SQFT 1183.60 SQFT Ground & Level 1 Ground, Level 1, Level 2 & Level 3 Ocean & Beach Ocean & Beach The above plans are subject to change as may be approved by the relevant authority. Areas are approximate measurements and subject to final survey. Plans are not to scale. 3 BEDROOM 3 BEDROOM TYPE C TYPE D (Penthouse) 214.53 SQM 177.29 SQM 2308.34 SQFT 1907.64 SQFT PANORAMIC PANORAMIC Level 2 Level 3 Ocean & Beach Ocean & Beach The above plans are subject to change as may be approved by the relevant authority. Areas are approximate measurements and subject to final survey. Plans are not to scale. Places of Interest Fort Frederick Harbour Built by the Portuguese in 1623 using the remains of the Koneswaram Trincomalee is the 2nd largest natural harbour in the world. Originally used Hindu Temple, it was rebuilt by the Dutch in 1665 and renamed as by the Chola army in 900AD, it was constantly fought over between the Sri Fort Frederick until the British took over in 1782. -

Sri Lanka Tourism: Poised for Growth
17 JUNE 2011 SRI LANKA TOURISM: POISED FOR GROWTH Inshita Wij Senior Associate www.hvs.com HVS India| 6th Floor, Building 8-C, DLF Cyber City Phase II, Gurgaon 122 002 INDIA Following the end of a three-decade long civil war in 2009, Sri Lanka has witnessed unprecedented growth. With a real GDP growth rate of 8% in 2010, a jump of 125.2% in the stock market in 2009, and 32% year-on-year growth in tourist arrivals in 2010, Sri Lanka is on its way to becoming a major tourism destination in South Asia. In the past one year, HVS India has received numerous inquiries about Sri Lanka from hotel operators, investors, and developers. These queries rightly come at a time when the country’s total rooms supply needs to be more than doubled in the next five years to meet the tourist arrivals targets. We have, therefore, in this article tried to highlight the current tourism landscape, highlighting the projected shortage of hotel rooms in the next five years and discussed in detail the factors that would help in tourism growth in the long term. We have also highlighted the future trends and challenges in the Sri Lankan hotel industry. The Current Tourism Landscape Sri Lanka witnessed a EXHIBIT 1: TOURIST ARRIVALS: 2000-2010 strong upsurge in tourism after the end of the civil 700,000 654,477 war in 2009. Tourism1, which forms 0.6% of the 600,000 549,308 total GDP of the country, 500,000 438,475 400,414 393,171 was one of the fastest 400,000 growing sectors in the 300,000 economy, growing by 200,000 39.8% in 2010 over 2009. -

Annual Performance Report of the District Secretariat
ANNUAL PERFORMANCE REPORT FOR THE YEAR 2019, TRINCOMALEE 뷒ස්ත්රි槊 ලේක කායාලය,ක臊ල臊ය,ි槔ණාමලය khtl;lr; nrayfk;, jpUNfhzkiy District Secretariat, Kachcheri, Trincomalee වාික කායස්ත්ාධන වාතාව tUlhe;j nraw;jpwd; mwpf;if ANNUAL PERFORMANCE REPORT 2019 0 ANNUAL PERFORMANCE REPORT FOR THE YEAR 2019, TRINCOMALEE Annual Performance Report for the year 2019 District Secretariat, Trincomalee Expenditure Head No 271 Contents Page Chapter 01 - Institutional Profile ………………………………………………………… 2-10 Chapter 02 – Progress and the Future Outlook …………………………………... 11 Chapter 03 - Overall Financial Performance for the Year ……………….……. 12-41 Chapter 04- Performance of the achieving Sustainable Development Goals (SDG) ………………………………………………. 42-45 Chapter 05 - Human Resource Profile ………………………………………………… 46-48 Chapter 06– Compliance Report ………………………………………………………… 49-54 1 ANNUAL PERFORMANCE REPORT FOR THE YEAR 2019, TRINCOMALEE Chapter 01 - Institutional Profile 1.1. Introduction Trincomalee District - A Glimpse The Boundary Trincomalee, a picturesque city with a natural 2arbor, scenic beauty, and military, commercial and historical importance, is situated in the eastern coast of Sri Lanka. Trincomalee District is boarded with Mulathivu District in North, Anuradhapura District in West and Polonnaruwa and Batticaloa Districts in the South. The History The history of Trincomalee goes back to a time of immemorial. The Mahavamsa & Chulavamsa, the two great chronicles, mention present Trincomalee as “Gokanna” , Gokarna, and “Gonagamaka” During the Anuradhapura and Polonnaruwa periods of island’s history. The Administration The Trincomalee District located in the center of Eastern Province covering an area of 2,727 square kilometers. The district is divided into 11 Divisional Secretary’s Divisions for administrative purpose. The DS Divisions are further sub-divided into 230 Grama Niladhari Divisions. -
Trincomalee District – 2007
BASIC POPULATION INFORMATION ON TRINCOMALEE DISTRICT – 2007 Preliminary Report Based on Special Enumeration – 2007 Department of Census and Statistics October 2007 ISBN 978-955-577-616-5 Foreword The Department of Census and Statistics (DCS), carried out a special enumeration in Eastern province and in Jaffna district in Northern province. The objective of this enumeration is to provide the necessary basic information needed to formulate development programmes and relief activities for the people. This preliminary publication for Trincomalee district has been compiled from the reports obtained from the District based on summaries prepared by enumerators and supervisors. A final detailed information will be disseminated after the computer processing of questionnaires. This preliminary release gives some basic information for Trincomalee district, such as population by divisional secretary’s division, urban/rural population, sex, age (under 18 years and 18 years and over) and ethnicity. Data on displaced persons due to conflict or tsunami are also included. Some important information which is useful for regional level planning purposes are given by Grama Niladhari Divisions. This enumeration is based on the usual residents of households in the district. These figures should be regarded as provisional. I wish to express my sincere thanks to the staff of the department and all other government officials and others who worked with dedication and diligence for the successful completion of the enumeration. I am also grateful to the general public for extending their fullest co‐operation in this important undertaking. This publication has been prepared by Population Census Division of this Department. D.B.P. Suranjana Vidyaratne Director General of Census and Statistics 10th October 2007 Department of Census and Statistics, 15/12, Maitland Crescent, Colombo 7. -

Tides of Violence: Mapping the Sri Lankan Conflict from 1983 to 2009 About the Public Interest Advocacy Centre
Tides of violence: mapping the Sri Lankan conflict from 1983 to 2009 About the Public Interest Advocacy Centre The Public Interest Advocacy Centre (PIAC) is an independent, non-profit legal centre based in Sydney. Established in 1982, PIAC tackles barriers to justice and fairness experienced by people who are vulnerable or facing disadvantage. We ensure basic rights are enjoyed across the community through legal assistance and strategic litigation, public policy development, communication and training. 2nd edition May 2019 Contact: Public Interest Advocacy Centre Level 5, 175 Liverpool St Sydney NSW 2000 Website: www.piac.asn.au Public Interest Advocacy Centre @PIACnews The Public Interest Advocacy Centre office is located on the land of the Gadigal of the Eora Nation. TIDES OF VIOLENCE: MAPPING THE SRI LANKAN CONFLICT FROM 1983 TO 2009 03 EXECUTIVE SUMMARY ....................................................................................................................... 09 Background to CMAP .............................................................................................................................................09 Report overview .......................................................................................................................................................09 Key violation patterns in each time period ......................................................................................................09 24 July 1983 – 28 July 1987 .................................................................................................................................10 -

Unspeakable Truth
This book is dedicated to the Tamils who perished waiting for justice Preface Contents This book traces the poignant history of Tamils in Sri Lanka after independence. It catalogues the Sri Lankan Tamils’ descent from a once thriving vibrant Nation to one Introduction that is today fi ghting for its very survival. This is a story about how a majority population consumed with religious chauvinism can corrupt a democratic process with untold 1. Documented genocide suffered by Tamils in Sri Lanka consequences. 1.1 State-aided Sinhala settlements in the Tamil homeland - Ethnic Cleansing 8 1.2 The Disenfranchisement of Tamils of Indian Origin 10 The book is organised into three sections covering the physical harm suffered by the 1.3 State-sponsored Riots against Tamils 12 Tamil community, the destruction of their cultural heritage and the attempts at negotiating 1.4 The 1983 Pogrom – a Watershed Event 16 a settlement which has come to nothing. The book also strikes a hopeful note at the 1.5 Progress from Pogroms to Aerial Bombings 20 end on how lasting peace can be achieved from the rubble of destruction. 1.6 The Torture and Murder of Civilians to win Submission 22 1.7 Rape as a Means of Suppression 26 The reader is likely to fi nd some images depicting examples of violence diffi cult and is 1.8 The Assassination of Political Leadership and Human Rights Activists 28 left to imagine the suffering endured by not only the victims but also their families and 1.9 Suppression and Violence against the Media 32 communities over the years. -

Luxury Beach Holiday 09 Nights
Luxury Beach Holiday 09 Nights Day 1 Airport – Polonnaruwa (approx: time 5 Hours 30 Mins) Overnight stay at Echo Lake House, Polonnaruwa Welcome & assist by Helanka Vacations, and proceed to Polonnaruwa. Visit Sightseeing in Polonnaruwa with visits to the largest man-made reservoir, the “Parakrama Samudraya” (the sea of Parakrama) built during King Parakrama’s reign. Also see the ruins of Shiva Devales and Hindu style decor which show the influence of Hindu culture on Sinhala Buddhist culture. The “Gal Vihara Complex” with its imposing Buddha sculptures in granite which is one of the most evocative pieces of sculpture in the world can be seen. Unique style of brick work architecture can also be seen at Lankathilaka Viharaya. The other interesting places are the ruins of the King’s Council Chamber, the Royal Pavilion, Kirivehera and the Gal Viaharaya. All these places show the height of stone craft and artistry of this period. Overnight stay at hotel in Echo Lake House. Day 2 Polonnaruwa – Pasikudah (approx: time 1 Hour 15 Mins) Overnight Stay at Anilana Pasikudah After breakfast proceed to Pasikudah. Pasikudah has a beautiful curved bay with extremely shallow water. Pasikudah is known to have one of the longest stretches of shallow coastline in the world. It is possible for people walk at least half a kilometer into the sea at certain points because the water is only a few inches deep and the current is relatively weak compared to the rest of Sri Lanka's coastline. Pasikudah Bay is a 2km long wide stretch of beach well protected by a shallow off shore reef. -

Humanitarian Operation Factual Analysis July 2006 – May 2009
HUMANITARIAN OPERATION FACTUAL ANALYSIS JULY 2006 – MAY 2009 MINISTRY OF DEFENCE DEMOCRATIC SOCIALIST REPUBLIC OF SRI LANKA HUMANITARIAN OPERATION FACTUAL ANALYSIS JULY 2006 – MAY 2009 MINISTRY OF DEFENCE JULY 2011 DEMOCRATIC SOCIALIST REPUBLIC OF SRI LANKA Humanitarian Operation—Factual Analysis TABLE OF CONTENTS Page I. EXECUTIVE SUMMARY 1 A. Overview of this Report 1 B. Overview of the Humanitarian Operation 1 PART ONE II. BACKGROUND 4 A. Overview of the LTTE 4 B. LTTE Atrocities against Civilians 6 C. Use of Child Soldiers by the LTTE 10 D. Ethnic Cleansing Carried out by the LTTE 10 E. Attacks on Democracy by the LTTE 11 F. The Global Threat posed by the LTTE 11 G. Proscription of the LTTE 12 III. SIZE AND SCOPE OF THE LTTE 13 A. Potency of the LTTE 13 B. Number of Cadres 14 C. Land Fighting Forces 14 D. The Sea Tiger Wing 17 E. The Air Tiger Wing 20 F. Black Tiger (Suicide) Wing 22 G. Intelligence Wing 22 H. Supply Network 23 I. International Support Mechanisms 25 J. International Criminal Network 27 – iii – Humanitarian Operation—Factual Analysis Page IV. GOVERNMENT EFFORTS FOR A NEGOTIATED SETTLEMENT 28 A. Overview 28 B. The Thimpu Talks – July to August 1985 29 C. The Indo-Lanka Accord – July 1987 30 D. Peace Talks – May 1989 to June 1990 32 E. Peace Talks – October 1994 to April 1995 33 F. Norwegian-Facilitated Peace Process – February 2002 to January 2008 35 G. LTTE Behaviour during 2002–2006 37 PART TWO V. RESUMPTION OF HOSTILITIES 43 VI. THE WANNI OPERATION 52 VII. -

12 Manogaran.Pdf
Ethnic Conflict and Reconciliation in Sri Lanka National Capilal District Boundarl3S * Province Boundaries Q 10 20 30 010;1)304050 Sri Lanka • Ethnic Conflict and Reconciliation in Sri Lanka CHELVADURAIMANOGARAN MW~1 UNIVERSITY OF HAWAII PRESS • HONOLULU - © 1987 University ofHawaii Press All Rights Reserved Manufactured in the United States ofAmerica Library ofCongress Cataloging-in-Publication-Data Manogaran, Chelvadurai, 1935- Ethnic conflict and reconciliation in Sri Lanka. Bibliography: p. Includes index. 1. Sri Lanka-Politics and government. 2. Sri Lanka -Ethnic relations. 3. Tamils-Sri Lanka-Politics and government. I. Title. DS489.8.M36 1987 954.9'303 87-16247 ISBN 0-8248-1116-X • The prosperity ofa nation does not descend from the sky. Nor does it emerge from its own accord from the earth. It depends upon the conduct ofthe people that constitute the nation. We must recognize that the country does not mean just the lifeless soil around us. The country consists ofa conglomeration ofpeople and it is what they make ofit. To rectify the world and put it on proper path, we have to first rec tify ourselves and our conduct.... At the present time, when we see all over the country confusion, fear and anxiety, each one in every home must con ., tribute his share ofcool, calm love to suppress the anger and fury. No governmental authority can sup press it as effectively and as quickly as you can by love and brotherliness. SATHYA SAl BABA - • Contents List ofTables IX List ofFigures Xl Preface X111 Introduction 1 CHAPTER I Sinhalese-Tamil -
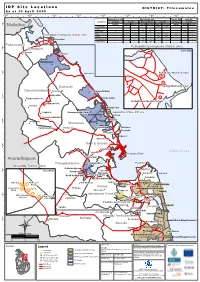
4A02e9560.Pdf
IDP Site Locations DISTRICT: Trincomalee As at 30 April 2009 80°50'0"E 80°55'0"E 81°0'0"E 81°5'0"E 81°10'0"E 81°15'0"E 81°20'0"E 81°25'0"E Displaced since 1983 Displaced after 2006 Tsunami IDPs Vanni IDPs Ds Division In camps In camps With host families Returnees In Camps In camps 9°0'0"N Families Families Individuals Families Individuals Families Individuals Families Families Individuals Mullaitivu Echchilampattai - - - - - 3,063 11,958 - - - Kinniya - - - 38 127 - - - - - Kuchchaveli 3 - - 26 95 - - - 1,745 5,666 Pulmoddai Emergency shelter sites Muttur - 645 2,067 - - 2,507 8,028 126 - - Seruvila - - - - - 136 512 - - Pulmoddai Thampalakamam - - - 17 43 - - - - - t[ Town & Gravets - - - 396 1,443 - - - - - Padavisripura 0.[ Pulmoddai Emergency shelter sites 8°55'0"N Shanthipura Inset Map Eramadu ! Thiriyai ! Kaddukulam ! Pulmoddai Muslim School 8°50'0"N Kuchaveli ! t[ t[ Kuchaveli Sinhala Mahavidyalaya Gomarankadawela Kivulekada Kumpurupiddi East ! ! ! Adampanai 8°45'0"N Pangurugaswewa Kumpurupiddi t[ ! ! Arafathnagar Muslim School ! Madawachchi Pettewa ! ! Valaiyoothu Tirappanai ! ! Kumpurupiddi South t[ Jaganathan Place IDP site Gopalapuram ! Nilaveli Gomarankadawala ! ! Veloor Mediyawa Nilaveli ! 8°40'0"N ! Morawewa ! ! Kambakkoddai Pankulam Periyakulam ! Mankanai ! ! ! Sempadu Sampaltivu ! ! !Uppuveli Kanniyai Town & Gravets 8°35'0"N !China Bay ! Indian Ocean Vayiriuttu ! TrincomaleeTown Kapalturai Anuradhapura ! Palampaddaru Foul Point Thampalakamam ! Kiliveddy Transit Sites ! Puthukudiyirippu Inset Map ! Kinniyai ! Muttur 8°30'0"N -

Eastern Province Biodiversity Profile and Conservation Action Plan
EASTERN PROVINCE BIODIVERSITY PROFILE AND CONSERVATION ACTION PLAN ,---~ ~- . f P-r~tP~tc\\ • ,_-- ··" : 9Hfct!O:"" 'l · . \ I -:: ~te, ·,t\.!~.' . ·· : -.;; ,,,;~(' \ J..,I ,.,. ~ , ,.. , ., Ht~,\~ _•! .. r 1t •.• \ Arc,-. <it::~:,~·--~:·.·:•·. .,~---·' i ( "oi· ... , --:-- ·- · ··-·· . ··· ···•• I . , . ., · - • , . .. ,., • ••·• • • • ' ' \ \ \ "\·•'-'"• •... ..... ' " ....,... t · · - · ~· ~ . ...... ~ .. .... .. i .'.\.\'•·,-··· :-·· ....... ........ ' Thangamuthu Jay~singam--· · ··· ···············-········ ; ~ • .... ... .r- l ~ ~~,· .. ~; ..~/ ~:,·1:u~ • •. __ .. ·-~ -..., ;\ai !I \ · . ~- l , ,i ,;,,~11·- • \ Vt '\!_;':!"!J ~ i • .• • ' l i"\.-~ lL -~:r,;.c-~(.;--- t • ~ • A Publication of Biodiversity Secretariat Ministry of Environment and Natural Resources Sri Lanka November 2008 ~~.. The Ministry of Environment and Natural Resources acknowledges the ADB/GEF, \.mM,L Government of the Netherlands funded Protected Area Management and Wildlife "'¥_. Conservation Project (Component "C") for providing financial assistance. Copyright: © Ministty of Environment and Natural Resources, Colombo, Sri Lanka Publisher: Biodiversity Secretariat Ministiy of Environment and Natural Resources, Sri Lanka Citation: Jayasingham, T. (2008) : Eastern Province Biodiversity Profile and Conservation Action Plan. The Biodiversity Secretariat, Ministry of Environment and Natural Resources, Sri Lanka. ISBN: 978-955-9120-92-6 First Print: December, 2008 Coordinated by: Lathika Hapuarachchi Saranga Jayasundara Printed by: Karunaratne & Sons (Pvt) Ltd.