Lankesteriana IV
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Lankesteriana IV
LANKESTERIANA 7(1-2): 229-239. 2007. DENSITY INDUCED RATES OF POLLINARIA REMOVAL AND DEPOSITION IN THE PURPLE ENAMEL-ORCHID, ELYTHRANTHERA BRUNONIS (ENDL.) A.S. GEORGE 1,10 2 3 RAYMOND L. TREMBLAY , RICHARD M. BATEMAN , ANDREW P. B ROWN , 4 5 6 7 MARC HACHADOURIAN , MICHAEL J. HUTCHINGS , SHELAGH KELL , HAROLD KOOPOWITZ , 8 9 CARLOS LEHNEBACH & DENNIS WIGHAM 1 Department of Biology, 100 Carr. 908, University of Puerto Rico – Humacao campus, Humacao, Puerto Rico, 00791-4300, USA 2 Natural History Museum, Cromwell Road, London SW7 5BD, UK 3 Department of Environment and Conservation, Species and Communities Branch, Locked Bag 104 Bentley Delivery Centre WA 6893, Australia 4 New York Botanic Garden, 112 Alpine Terrace, Hilldale, NJ 00642, USA 5 School of Life Sciences, University of Sussex, Falmer, Brighton, Sussex, BN1 9QG, UK 6 IUCN/SSC Orchid Specialist Group Secretariat, 36 Broad Street, Lyme Regis, Dorset, DT7 3QF, UK 7 University of California, Ecology and Evolutionary Biology, Irvine, CA 92697, USA 8 Massey University, Allan Wilson Center for Molecular Ecology and Evolution 9 Smithsonian Institution, Smithsonian Environmental Research Center, Box 28, Edgewater, MD 21037, USA 10 Author for correspondence: [email protected] RESUMEN. La distribución y densidad de los individuos dentro de las poblaciones de plantas pueden afectar el éxito reproductivo de sus integrantes. Luego de describir la filogenia de las orquideas del grupo de las Caladeniideas y su biología reproductiva, evaluamos el efecto de la densidad en el éxito reproductivo de la orquídea terrestre Elythranthera brunonis, endémica de Australia del Oeste. El éxito reproductivo de esta orquídea, medido como la deposición y remoción de polinios, fue evaluado. -

South East Flora
Regional Species Conservation Assessments DENR South East Region Complete Dataset for all Flora Assessments Dec 2011 In Alphabetical Order of Species Name MAP ID FAMILY NAME PLANT FORM NSX CODE SPECIES NAME COMMON NAME SOUTH EAST Regional EAST SOUTH Status Regional EAST SOUTH Status Score Regional Trend EAST SOUTH Score Regional EAST SOUTH Status+Trend Score SOUTH EAST Regional Trend EAST SOUTH FAMILY FAMILY NUMBER (CENSUS OF SA) EPBCACTSTATUSCODE NPWACTSTATUSCODE LASTOBSERVED_in_SE TOTAL_in_SA TOTAL_in_SE %_SOUTH_EAST_REGION EofO_in_SE_All_km2 EofO_in_SE_Recent_km2 AofO_in_SE_All_km2 AofO_in_SE_Recent_km2 711 91.182 LEGUMINOSAE legumes Y01536 Acacia acinacea Wreath Wattle 2009 814 60 7.37 3000 1700 48 27 LC 1 0 0.3 1.3 712 91.182 LEGUMINOSAE legumes K01545 Acacia brachybotrya Grey Mulga-bush 2001 563 18 3.20 800 500 16 9 RA 3 0 0.3 3.3 713 91.182 LEGUMINOSAE legumes M01554 Acacia continua Thorn Wattle 1974 836 1 0.12 100 1 VU 4 DD 0.0 4.0 714 91.182 LEGUMINOSAE legumes C05237 Acacia cupularis Cup Wattle 2002 577 83 14.38 4700 1500 65 20 LC 1 0 0.3 1.3 716 91.182 LEGUMINOSAE legumes K01561 Acacia dodonaeifolia Hop-bush Wattle R 2002 237 33 13.92 800 400 19 6 RA 3 0 0.3 3.3 718 91.182 LEGUMINOSAE legumes M01562 Acacia enterocarpa Jumping-jack Wattle EN E 2008 92 16 17.39 700 400 10 7 VU 4 0 0.3 4.3 719 91.182 LEGUMINOSAE legumes C05985 Acacia euthycarpa Wallowa 1992 681 7 1.03 500 100 7 1 RA 3 - 0.4 3.4 720 91.182 LEGUMINOSAE legumes S01565 Acacia farinosa Mealy Wattle 1997 325 88 27.08 4000 1600 65 23 NT 2 0 0.3 2.3 721 91.182 LEGUMINOSAE -

AUSTRALIAN ORCHID NAME INDEX (27/4/2006) by Mark A. Clements
AUSTRALIAN ORCHID NAME INDEX (27/4/2006) by Mark A. Clements and David L. Jones Centre for Plant Biodiversity Research/Australian National Herbarium GPO Box 1600 Canberra ACT 2601 Australia Corresponding author: [email protected] INTRODUCTION The Australian Orchid Name Index (AONI) provides the currently accepted scientific names, together with their synonyms, of all Australian orchids including those in external territories. The appropriate scientific name for each orchid taxon is based on data published in the scientific or historical literature, and/or from study of the relevant type specimens or illustrations and study of taxa as herbarium specimens, in the field or in the living state. Structure of the index: Genera and species are listed alphabetically. Accepted names for taxa are in bold, followed by the author(s), place and date of publication, details of the type(s), including where it is held and assessment of its status. The institution(s) where type specimen(s) are housed are recorded using the international codes for Herbaria (Appendix 1) as listed in Holmgren et al’s Index Herbariorum (1981) continuously updated, see [http://sciweb.nybg.org/science2/IndexHerbariorum.asp]. Citation of authors follows Brummit & Powell (1992) Authors of Plant Names; for book abbreviations, the standard is Taxonomic Literature, 2nd edn. (Stafleu & Cowan 1976-88; supplements, 1992-2000); and periodicals are abbreviated according to B-P-H/S (Bridson, 1992) [http://www.ipni.org/index.html]. Synonyms are provided with relevant information on place of publication and details of the type(s). They are indented and listed in chronological order under the accepted taxon name. -

Flowering Times of Tasmanian Orchids: a Practical Guide for Field Botanists
Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists Mark Wapstra (Environmental Consulting Options Tasmania, ECOtas) 4th Edition, July 2018 Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists 4th Edition (July 2018) MARK WAPSTRA Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists FOREWORD TO FIRST EDITION (2008) This document fills a significant gap in the Tasmanian orchid literature. Given the inherent difficulties in locating and surveying orchids in their natural habitat, an accurate guide to their flowering times will be an invaluable tool to field botanists, consultants and orchid enthusiasts alike. Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists has been developed by Tasmania’s leading orchid experts, drawing collectively on many decades of field experience. The result is the most comprehensive State reference on orchid flowering available. By virtue of its ease of use, accessibility and identification of accurate windows for locating our often-cryptic orchids, it will actually assist in conservation by enabling land managers and consultants to more easily comply with the survey requirements of a range of land-use planning processes. The use of this guide will enhance efforts to locate new populations and increase our understanding of the distribution of orchid species. The Threatened Species Section commends this guide and strongly recommends its use as a reference whenever surveys for orchids are undertaken. Matthew Larcombe Project Officer (Threatened Orchid and Euphrasia) Threatened Species Section, Department of Primary Industries, Parks, Water & Environment March 2008 DOCUMENT AVAILABILITY This document is freely available as a PDF file downloadable from the following websites: www.fpa.tas.gov.au; www.dpipwe.tas.gov.au; www.ecotas.com.au. -

Flowering Times of Tasmanian Orchids: a Practical Guide for Field Botanists
Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists 0 Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists FOREWORD This document fills a significant gap in the Tasmanian orchid literature. Given the inherent difficulties in locating and surveying orchids in their natural habitat, an accurate guide to their flowering times will be an invaluable tool to field botanists, consultants and orchid enthusiasts alike. Flowering Times of Tasmanian Orchids: A Practical Guide for Field Botanists has been developed by Tasmania’s leading orchid experts, drawing collectively on many decades of field experience. The result is the most comprehensive State reference on orchid flowering available. By virtue of its ease of use, accessibility and identification of accurate windows for locating our often-cryptic orchids, it will actually assist in conservation by enabling land managers and consultants to more easily comply with the survey requirements of a range of land-use planning processes. The use of this guide will enhance efforts to locate new populations and increase our understanding of the distribution of orchid species. The Threatened Species Section commends this guide and strongly recommends its use as a reference whenever surveys for orchids are undertaken. Matthew Larcombe Project Officer (Threatened Orchid and Euphrasia) Threatened Species Section, Department of Primary Industries, Parks, Water & Environment March 2008 DOCUMENT AVAILABILITY This document is available as a PDF file downloadable from the following websites: www.fpa.tas.gov.au www.dpipwe.tas.gov.au www.ecotas.com.au It may also be requested directly from the authors (see contact details below). -

Biodiversity and Endemism Within the Mount Canobolas Volcanic Complex
Biodiversity and Endemism within the Mount Canobolas Volcanic Complex R������ W. M���1 and C���� C. B����2 Published on 23 December 2019 at https://openjournals.library.sydney.edu.au/index.php/LIN/index 1593 Cargo Road, Orange NSW 2800 ([email protected]); 2FloraSearch, PO Box 300, Orange NSW 2800 ([email protected]) Medd, R.W. and Bower, C.C. (2019). Biodiversity and endemism within the Mount Canobolas volcanic complex. Proceedings of the Linnean Society of New South Wales 141, S45-S83. Mt Canobolas State Conservation Area (SCA) hosts a small remnant of sub-alpine vegetation consisting of seven recognisable communities with the heathlands on the rock plates appearing to be unique to the SCA. The SCA has a known biota of 884 native species that includes 14 threatened species and at least 10 endemic taxa. Some 200 species are regionally significant, being either rare or at the limits of known geographic range. The vascular flora is particularly species-rich being considerably more diverse than nearby regional reserves and over 12 fold richer than comparable areas of the Kosciusko National Park. One of three endangered ecological communities, the Mt Canobolas Xanthoparmelia Lichen Community, is unique to the volcanic province. While there is some indication the endemic lithophytic lichens, the threatened Eucalyptus canobolensis and the heath communities may be substrate specific, there is no strong evidence of a geological association among other flora and fauna. We postulate that the presence of multiple endemic species reflects the geographic isolation which has provided an environment for species evolution by vicariance. Alternatively, Mt Canobolas has acted as a refugium for formerly widespread species that have become extinct elsewhere. -
Checklist of the Orchids of Australia Including Its Island Territories
Checklist of the Orchids of Australia Including its Island Territories Gary N. Backhouse Robert J. Bates Andrew P. Brown Lachlan M. Copeland Second Edition Contents Introduction ........................................................................................ page 1 Australia orchid genera ...................................................................... page 4 Australia species ................................................................................. page 7 Non-native (exotic or weed) species ................................................ page 57 Australia hybrids ............................................................................... page 58 Australian Capital Territory species .................................................. page 69 Australian Capital Territory hybrids .................................................. page 72 New South Wales species ................................................................. page 73 New South Wales hybrids ................................................................. page 86 Northern Territory species ............................................................... page 88 Queensland species .......................................................................... page 89 Queensland hybrids .......................................................................... page 99 South Australia species ................................................................... page 100 South Australia hybrids .................................................................. -
KONGJAROON Chutima-Thesis Nosignature.Pdf
Investigation into the Use of Molecular Methods to Distinguish between Species of Caladenia Subgenus Calonema Chutima Kongjaroon A thesis submitted in a fulfillment of the requirements for the degree of Doctor of Philosophy School of Engineering and Sciences Faculty of Health, Engineering and Science Victoria University Melbourne Australia 2012 Thesis abstract The phylogeny of typical spider orchid (Caladenia subgenus Calonema) is investigated for the first time. The analyses were performed using 17 RAPD and 10 ISSR primers on 30 taxa representing the three spider orchid groups (the dilatata, patersonii and reticulata groups) yielding 135 RAPD and 63 ISSR polymorphic markers. The average number of polymorphic markers produced from 17 RAPD and 10 ISSR primers were 5.12 and 4.48, respectively. 76 RAPD markers and 38 ISSR markers were polymorphic within spider orchid species. The highest number of amplified DNA fragments were produced from OPE15 (8.77 fragments) and UBC 842 (6.71 fragments) while OPF04 (2.93 fragments) and UBC 825 (3.02 fragments) gave the smallest number of amplification products. The average Dice genetic similarity of pairs of individuals within a species ranged from 0.772 to 0.939 based on RAPD and from 0.770 to 0.976 for ISSR data. The 117 spider orchid individuals analysed by RAPD, ISSR and combined data using cluster analysis were classified into three groups that correlate with those based on morphological analysis: reticulata, patersonii and dilatata. The phylogenetic analysis based on three data sets, presented relationships at the intraspecific level that were well supported by bootstrapping. However, the interspecific relationships within the groups remained unclear. -
Part 10 ESP Intro
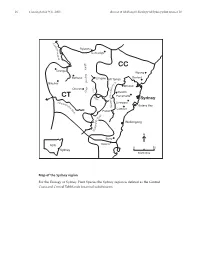
16 Cunninghamia 9(1): 2005 Benson & McDougall, Ecology of Sydney plant species 10 M a c q u Rylstone a r i e Coricudgy R i v e r e g n CC a Orange R Wyong g n i Gosford Bathurst d i Lithgow v Mt Tomah i Blayney D R. y r Windsor C t u a o b Oberon s e x r e s G k Penrith w a R Parramatta CT H i ve – Sydney r n a Abe e Liverpool rcro p m e b Botany Bay ie N R Camden iv Picton er er iv R y l l i Wollongong d n o l l o W N Berry NSW Nowra 050 Sydney kilometres Map of the Sydney region For the Ecology of Sydney Plant Species the Sydney region is defined as the Central Coast and Central Tablelands botanical subdivisions. Cunninghamia 9(1): 2005 Benson & McDougall, Ecology of Sydney plant species 10 17 Ecology of Sydney plant species Part 10 Monocotyledon families Lemnaceae to Zosteraceae Doug Benson and Lyn McDougall Royal Botanic Gardens and Domain Trust, Sydney, AUSTRALIA 2000. Email: [email protected] Abstract: Ecological data in tabular form are provided on 668 plant species of the families Lemnaceae to Zosteraceae, 505 native and 163 exotics, occurring in the Sydney region, defined by the Central Coast and Central Tablelands botanical subdivisions of New South Wales (approximately bounded by Lake Macquarie, Orange, Crookwell and Nowra). Relevant Local Government Areas are Auburn, Ashfield, Bankstown, Bathurst, Baulkham Hills, Blacktown, Blayney, Blue Mountains, Botany, Burwood, Cabonne, Camden, Campbelltown, Canada Bay, Canterbury, Cessnock, Crookwell, Evans, Fairfield, Greater Lithgow, Gosford, Hawkesbury, Holroyd, Hornsby, Hunters Hill, Hurstville, Kiama, Kogarah, Ku-ring-gai, Lake Macquarie, Lane Cove, Leichhardt, Liverpool, Manly, Marrickville, Mosman, Mulwaree, North Sydney, Oberon, Orange, Parramatta, Penrith, Pittwater, Randwick, Rockdale, Ryde, Rylstone, Shellharbour, Shoalhaven, Singleton, South Sydney, Strathfield, Sutherland, Sydney City, Warringah, Waverley, Willoughby, Wingecarribee, Wollondilly, Wollongong, Woollahra and Wyong. -

2019 Proposed Amendments to the Status of Threatened Species in South Australia
2019 Proposed Amendments to the Status of Threatened Species in South Australia Proposed amendments to the Threatened Species Schedules of the South Australian National Parks and Wildlife Act 1972 In South Australia, threatened plants and animals are listed in schedules of the National Parks and Wildlife Act 1972 (NPW Act), to identify and protect species of conservation concern. The “Threatened Species Schedules” of the NPW Act currently provide for three categories of threat: Endangered (Schedule 7), Vulnerable (Schedule 8) and Rare (Schedule 9), which are designed to reflect the relative risk of extinction in the wild. With the assistance of scientific experts, the Department for Environment and Water assesses the conservation status of South Australia’s plant and animal species. Based on these assessments, the Threatened Species Schedules of the NPW Act are periodically amended to reflect new information, taxonomic changes, and changes in the threat status of species. Public comment is now being sought on recommended changes (new listings, delisting or changes in threat category) for 65 species, to inform proposed amendments to the Threatened Species Schedules. Comments relating to the eligibility of these species for the recommended status classifications and criteria, together with additional information relevant to the assessments, should be sent to [email protected]. Comments will be accepted until 16 August 2019. Suggestions for priority assessments of species not listed here will also be considered for inclusion in the ongoing assessments work plan. Assessments process The South Australian Department for Environment and Water assesses the conservation status of the State’s plant and animal species with considerable assistance from panels of experts on different taxonomic groups. -

Vascular Flora of the South East Forests Region, Eden, New South Wales
219 Vascular Flora of the South East Forests region, Eden, New South Wales David A. Keith, Jackie Miles and Berin D. E. Mackenzie Abstract Keith, David A.1, Miles, Jackie2, and Mackenzie, Berin, D. E.1 (1NSW National Parks and Wildlife Service, PO Box 1967, Hurstville, NSW, Australia 2220; 2Hawkshead Road, Brogo, NSW, Australia 2550) 1999. Vascular flora of the South East Forests region, Eden, New South Wales. Cunninghamia 6(1): 219–281. An exhaustive review of available information revealed 1522 native vascular plant taxa with confirmed records in the Eden region. A further 136 taxa potentially occur in the region, but these remain to be confirmed. Families represented by large numbers of taxa in the indigenous flora include the Fabaceae, Poaceae, Asteraceae, Myrtaceae and Orchidaceae. Species frequencies were highly skewed with 74% of the flora represented in less than 1% of survey samples and only three taxa represented in more than 40% of samples. A large number of records (350 taxa) were evaluated and refuted, emphasising the importance of critical validation when compiling biological inventory data from multiple sources. Errors are most likely to arise from nomenclatural synonymy and other duplications, misidentifications and erroneous locality information. The confirmed vascular flora of the Eden region is relatively species-rich compared with other regions in tropical, temperate and semi-arid Australia, although these regions are less well-known botanically. However, the proportion of endemism (1%) is comparatively low, with 18 taxa unique to the region and a further 14 taxa with distributions extending just beyond the region. The Eden region includes 32 taxa listed as endangered or vulnerable in Australia and a further six taxa listed as endangered in New South Wales but not throughout Australia. -

Australian Orchid Name Index (13/6/2008)
AUSTRALIAN ORCHID NAME INDEX (13/6/2008) by Mark A. Clements and David L. Jones Centre for Plant Biodiversity Research/Australian National Herbarium GPO Box 1600 Canberra ACT 2601 Australia Corresponding author: [email protected] INTRODUCTION The Australian Orchid Name Index (AONI) provides the currently accepted scientific names, together with their synonyms, of all Australian orchids including those in external territories. The appropriate scientific name for each orchid taxon is based on data published in the scientific or historical literature, and/or from study of the relevant type specimens or illustrations and study of taxa as herbarium specimens, in the field or in the living state. Structure of the index: Genera and species are listed alphabetically. Accepted names for taxa are in bold, followed by the author(s), place and date of publication, details of the type(s), including where it is held and assessment of its status. The institution(s) where type specimen(s) are housed are recorded using the international codes for Herbaria (Appendix 1) as listed in Holmgren et al’s Index Herbariorum (1981) continuously updated, see [http://sciweb.nybg.org/science2/IndexHerbariorum.asp]. Citation of authors follows Brummit & Powell (1992) Authors of Plant Names; for book abbreviations, the standard is Taxonomic Literature, 2nd edn. (Stafleu & Cowan 1976-88; supplements, 1992-2000); and periodicals are abbreviated according to B-P- H/S (Bridson, 1992) [http://www.ipni.org/index.html]. Synonyms are provided with relevant information on place of publication and details of the type(s). They are indented and listed in chronological order under the accepted taxon name.