Μεταπτυχιακή Μελέτη: «Ποικιλότητα Ειδών Αυχενορρύγχων (Auchenorrhyncha) Σε Καλλιέργειες Μηδικής»
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Topic Paper Chilterns Beechwoods
. O O o . 0 O . 0 . O Shoping growth in Docorum Appendices for Topic Paper for the Chilterns Beechwoods SAC A summary/overview of available evidence BOROUGH Dacorum Local Plan (2020-2038) Emerging Strategy for Growth COUNCIL November 2020 Appendices Natural England reports 5 Chilterns Beechwoods Special Area of Conservation 6 Appendix 1: Citation for Chilterns Beechwoods Special Area of Conservation (SAC) 7 Appendix 2: Chilterns Beechwoods SAC Features Matrix 9 Appendix 3: European Site Conservation Objectives for Chilterns Beechwoods Special Area of Conservation Site Code: UK0012724 11 Appendix 4: Site Improvement Plan for Chilterns Beechwoods SAC, 2015 13 Ashridge Commons and Woods SSSI 27 Appendix 5: Ashridge Commons and Woods SSSI citation 28 Appendix 6: Condition summary from Natural England’s website for Ashridge Commons and Woods SSSI 31 Appendix 7: Condition Assessment from Natural England’s website for Ashridge Commons and Woods SSSI 33 Appendix 8: Operations likely to damage the special interest features at Ashridge Commons and Woods, SSSI, Hertfordshire/Buckinghamshire 38 Appendix 9: Views About Management: A statement of English Nature’s views about the management of Ashridge Commons and Woods Site of Special Scientific Interest (SSSI), 2003 40 Tring Woodlands SSSI 44 Appendix 10: Tring Woodlands SSSI citation 45 Appendix 11: Condition summary from Natural England’s website for Tring Woodlands SSSI 48 Appendix 12: Condition Assessment from Natural England’s website for Tring Woodlands SSSI 51 Appendix 13: Operations likely to damage the special interest features at Tring Woodlands SSSI 53 Appendix 14: Views About Management: A statement of English Nature’s views about the management of Tring Woodlands Site of Special Scientific Interest (SSSI), 2003. -

Additional Notes on the Some Aphrophorid Spittlebugs of Eastern Anatolia (Hemiptera: Cercopoidea: Aphrophoridae)*
I. Ozgen 1 et al. ISSN 2587-1943 ADDITIONAL NOTES ON THE SOME APHROPHORID SPITTLEBUGS OF EASTERN ANATOLIA (HEMIPTERA: CERCOPOIDEA: APHROPHORIDAE)* İnanç Özgen 1, Aykut Topdemir 2, Fariba Mozaffarian 3 scientific note The study was carried out to determine Aphrophoridae species in Eastern Anatolia in 2018. Five species were collected by sweeping net on herbs. The collected specimens were identified as: Aphrophora salicina (Goeze, 1778), Lepyronia coleoptrata (Linnaeus, 1758), Paraphilaenus notatus (Mulsant & Rey, 1855), Philaenus spumarius (Linnaeus, 1758) and Neophilaenus campestris (Fallén, 1805). The species P. spumarius and L. coleoptrata were the most abundant species and the others were rather rare. The species of family Aphrophoridae are xylem feeders so they are considered as candidates for transmitting bacteria Xylella fastidiosa. Therefore, the role of the identified species in the agricultural ecosystems in the collecting sites needs to be studied. Key words: Hemiptera, Aphrophoridae, Fauna, Eastern Anatolia 1 Introduction The Aphrophoridae or spittlebugs are a family of Note: N. campestris prefer mostly grasslands, insects belonging to the order Hemiptera. Nymphs of Neophilaenus campestris Fallén showed harbour the Aphrophoridae secrete a frothy saliva-like mass, which bacterium in their body (Elbeaino et al.,2014; Moussa et al., gives the name “spittlebugs” for insects in the superfamily. 2017). The species of family Aphrophoridae are xylem feeders so Paraphilaenus notatus (Mulsant & Rey, 1855), they are considered as candidates for transmitting bacteria Xylella fastidiosa. In this study were carried out to Material examined: Elazığ, Aşağı çakmak village, determine of Aphrophorid fauna in Eastern Anatolia of 18.V.2018, 6 exs. Turkey. Note: It was determined to potential vector of Xylella 2 Material and Method fastidiosa. -

Charles R. Bartlett & Gernot Kunz (2015) A
Zootaxa 3963 (4): 598–600 ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Erratum ZOOTAXA Copyright © 2015 Magnolia Press ISSN 1175-5334 (online edition) http://dx.doi.org/10.11646/zootaxa.3963.4.7 http://zoobank.org/urn:lsid:zoobank.org:pub:374DEA43-B853-4291-8CB9-17C59DE8BDDB CHARLES R. BARTLETT & GERNOT KUNZ (2015) A new genus and species of delphacid planthopper (Hemiptera: Fulgoroidea: Delphacidae) from Central America with a preliminary regional species list. Zootaxa, 3946(4): 510–518. Table 1 was inadvertently omitted from the text. It is provided as follows. Table 1. List of delphacid species found in Costa Rica and adjacent countries (L = literature record, S = specimen record, E = error). Costa Species Nicaragua Rica Panama References and Comments Delphacidae Asiracinae: Asiracini Metcalf 1943, Maes & O’Brien 1988, Maes & Tellez Robleto 1988, Bartlett Copicerus irroratus Swartz, 1802 L, S L, S L, S et al. 2014 Asiracinae: Idiosystanini Metcalf 1943, Maes & Tellez Robleto 1988, Hedrick-Zeller & Wilson 2010; Pentagramma bivittata Crawford, 1914 L, S L, S Bartlett et al. 2014 Asiracinae: Tetrasteirini Tetrasteira solata Barringer & Bartlett, 2011 L, S Barringer & Bartlett 2011 Tetrasteira trimaculata Barringer & Bartlett 2011 L, S L, S Barringer & Bartlett 2011 Asiracinae: Ugyopini Ugyops brunneus (Fowler, 1905) L Fowler 1905, Metcalf 1943 Ugyops godmani (Fowler, 1905) L L, S Fowler 1905, Metcalf 1943 Ugyops palliatus Fennah, 1964 L Fennah 1964 Ugyops stigmatus (Crawford, 1914) L, S Crawford, 1914, Metcalf 1938, 1943 Ugyops sp. S Plesiodelphacinae Crawford, 1914, Maes & O’Brien Burnilia pictifrons (Stål, 1864) L 1988 Burnilia n. sp. S Delphacinae: Saccharosydnini Neomalaxa flava Muir, 1918 S S Maes & O’Brien 1988, Maes & Saccharosydne saccharivora Tellez Robleto 1988, Bartlett et al. -

Biodiversity Climate Change Impacts Report Card Technical Paper 12. the Impact of Climate Change on Biological Phenology In
Sparks Pheno logy Biodiversity Report Card paper 12 2015 Biodiversity Climate Change impacts report card technical paper 12. The impact of climate change on biological phenology in the UK Tim Sparks1 & Humphrey Crick2 1 Faculty of Engineering and Computing, Coventry University, Priory Street, Coventry, CV1 5FB 2 Natural England, Eastbrook, Shaftesbury Road, Cambridge, CB2 8DR Email: [email protected]; [email protected] 1 Sparks Pheno logy Biodiversity Report Card paper 12 2015 Executive summary Phenology can be described as the study of the timing of recurring natural events. The UK has a long history of phenological recording, particularly of first and last dates, but systematic national recording schemes are able to provide information on the distributions of events. The majority of data concern spring phenology, autumn phenology is relatively under-recorded. The UK is not usually water-limited in spring and therefore the major driver of the timing of life cycles (phenology) in the UK is temperature [H]. Phenological responses to temperature vary between species [H] but climate change remains the major driver of changed phenology [M]. For some species, other factors may also be important, such as soil biota, nutrients and daylength [M]. Wherever data is collected the majority of evidence suggests that spring events have advanced [H]. Thus, data show advances in the timing of bird spring migration [H], short distance migrants responding more than long-distance migrants [H], of egg laying in birds [H], in the flowering and leafing of plants[H] (although annual species may be more responsive than perennial species [L]), in the emergence dates of various invertebrates (butterflies [H], moths [M], aphids [H], dragonflies [M], hoverflies [L], carabid beetles [M]), in the migration [M] and breeding [M] of amphibians, in the fruiting of spring fungi [M], in freshwater fish migration [L] and spawning [L], in freshwater plankton [M], in the breeding activity among ruminant mammals [L] and the questing behaviour of ticks [L]. -

Hemiptera: Cicadellidae and Delphacidae) in Brazilian Maize Crops
1470 Florida Entomologist 96(4) December 2013 ABUNDANCE AND SPECIES RICHNESS OF LEAFHOPPERS AND PLANTHOPPERS (HEMIPTERA: CICADELLIDAE AND DELPHACIDAE) IN BRAZILIAN MAIZE CROPS 1 2 2 3 CHARLES MARTINS DE OLIVEIRA , ELIZABETH DE OLIVEIRA , ISABEL REGINA PRAZERES DE SOUZA , ELCIO ALVES , 4 5 5 6 WILLIAM DOLEZAL , SUSANA PARADELL , ANA MARIA MARINO DE REMES LENICOV AND MARINA REGINA FRIZZAS 1Embrapa Cerrados. C.P. 08223, Planaltina, Brasília/DF, 73310-970, Brazil 2Embrapa Milho e Sorgo. C. P. 151, Sete Lagoas/MG, 35701970, Brazil 3DuPont do Brazil S.A, Divisão Pioneer Sementes. C.P. 1014, Itumbiara/GO, 75503971, Brazil 4Pioneer Hi-Bred International, Inc. C.P. 1014, Itumbiara/GO, 75503971, Brazil 5Facultad de Ciencias Naturales y Museo, División Entomologia, Universidad Nacional de La Plata. Paseo del Bosque s/n – La Plata (1900), Buenos Aires, Argentina 6Universidade de Brasília, Departamento de Zoologia, Instituto de Ciências Biológicas, Brasília/DF, Brazil, 70910-900 Corresponding author; E-mail: [email protected] ABSTRACT Insects in the Cicadellidae and Delphacidae families, common in grasses, are an important group of vectors of viruses and mollicutes, which cause diseases in several plant species. The goal of this study was to evaluate the abundance and species richness of Cicadellidae and Delphacidae and the presence of potential vectors of viruses and mollicutes in maize crops in Brazil. Insects were collected using sweep nets in maize crops in 48 counties of 8 states, distributed in 4 regions of Brazil in the yr 2005, 2006 and 2007, with a total of 198 samples. The collected material was screened, and the leafhoppers and planthoppers were identi- fied at the species level. -

Durham E-Theses
Durham E-Theses Studies on the auchenorrhyncha (hemoptera insecta) of Pennine moorland with special reference to the ceropidae Whittaker, John B. How to cite: Whittaker, John B. (1963) Studies on the auchenorrhyncha (hemoptera insecta) of Pennine moorland with special reference to the ceropidae, Durham theses, Durham University. Available at Durham E-Theses Online: http://etheses.dur.ac.uk/10475/ Use policy The full-text may be used and/or reproduced, and given to third parties in any format or medium, without prior permission or charge, for personal research or study, educational, or not-for-prot purposes provided that: • a full bibliographic reference is made to the original source • a link is made to the metadata record in Durham E-Theses • the full-text is not changed in any way The full-text must not be sold in any format or medium without the formal permission of the copyright holders. Please consult the full Durham E-Theses policy for further details. Academic Support Oce, Durham University, University Oce, Old Elvet, Durham DH1 3HP e-mail: [email protected] Tel: +44 0191 334 6107 http://etheses.dur.ac.uk 2 ABSTRACT Studies on the Auchenorrhyncha (Heina>ptera - insecta) of Pennine Moorland with special reference to the Cercopidae. Notes on the autecology of 32 species of Auchenorrhyncha from the Moor House National Nature Reserve are given. Studies were made of the microclimates of the common vegetation types on which these occur and it is shown that the size and function as a temperature regulator of the spittle (produced by nymphs of the Cercopidae) is associated with these gradients. -

Two New Leafhopper Genera of the Alebroides Genus Group (Hemiptera: Cicadellidae: Typhlocybinae) from China, with a Key to Genera of the Group
Entomological Science (2017) doi: 10.1111/ens.12260 ORIGINAL ARTICLE Two new leafhopper genera of the Alebroides genus group (Hemiptera: Cicadellidae: Typhlocybinae) from China, with a key to genera of the group Ye XU1,SihanLU1, Yuru WANG1, Christopher H. DIETRICH2 and Daozheng QIN1 1 Key Laboratory of Plant Protection Resources and Pest Management of the Ministry of Education, Entomological Museum, Northwest A&F University, Yangling, China and 2 Illinois Natural History Survey, Prairie Research Institute, University of Illinois, Champaign, Illinois, USA Abstract Two new microleafhopper genera in the Alebroides genus group, Nulliata Lu, Xu & Qin, gen. nov., based on the type species N. rubrostriata Lu, Xu & Qin, sp. nov.,andInflatopina Lu, Dietrich & Qin, gen. nov., based on the type species I. intonsa Lu, Dietrich & Qin, sp. nov., are described from southwest China. Five known species in the Alebroides sohii species group are transferred to Inflatopina as new combinations. Keys to genera of the Alebroides genus group and species of Inflatopina are given. Key words: Auchenorrhyncha, microleafhopper, morphology, taxonomy. INTRODUCTION the cosmopolitan genus Empoasca Walsh, 1862). In contrast, the Alebroides group appears more stable in The microleafhopper tribe Empoascini comprises more classification, comprising 156 species in 25 genera so than 1,000 species in 85 previously described genera far, widely distributed in the Oriental, Palaearctic, from throughout the world. It can be distinguished from Afrotropical and Australian Regions but absent from other tribes of Typhlocybinae as follows: ocelli usually the New World (Xu et al. 2016). well developed; forewing without appendix; hind wing In China, the Alebroides group includes 14 genera and submarginal vein extended between apices of veins 58 species known to date (see checklist). -

Tracking Vectors of Bacteria and Phytoplasmas Threatening Europe’S Major Crops (VECTRACROP)
Euphresco Final Report Tracking vectors of bacteria and phytoplasmas threatening Europe’s major crops (VECTRACROP) Topic area Phloem and xylem feeding insect vectors, fruit and field crops, bacteria and phytoplasmas of phytosanitary concern - Topic Description 2015-D-168 Topic title Tracking vectors of bacteria and phytoplasmas threatening Europe’s major crops (VECTRACROP) 1. Administrative Details . Applicant / Coordinator – Partner 1 Organisation Institute for AgriculturaI and Fisheries Research - ILVO Name of contact Kris De Jonghe, Ph.D. Gender: M (incl. Title) Postal address Burg. Van Gansberghelaan 96, B- 9820 Merelbeke, Belgium E-mail [email protected]; [email protected] Phone ++32 9/ 272 24 48 Applicant – Partner 2 Organisation CRA-W Name of contact Thibaut Olivier, Ir Gender: M (incl. Title) Département Sciences du Vivant (CRAW), Unité Biologie des Postal address nuisibles et Biovigilance, Bâtiment Marchal, Rue de Liroux 4, B- 5030 Gembloux, Belgium E-mail [email protected] Phone ++32 81/ 62 03 39 Applicant – Partner 3 Organisation ANSES Name of contact Reynaud Philippe, Ph.D. Gender: M (incl. Title) Anses Laboratoire de la Santé des Végétaux [Plant Health Laboratory] Postal address 755 avenue du campus Agropolis CS 30016 FR-34988 Montferrier-sur-Lez Cedex E-mail [email protected] Phone + 33 (0)4 67 02 25 10 Applicant – Partner 4 Organisation INIAV Name of contact Célia Mateus- Researcher, Ph.D.; Esmeraldina Gender F (incl. Title) Sousa- Researcher, Ph.D. : Av. da República, Quinta do Marquês Postal address 2780-157 Oeiras – Portugal E-mail [email protected]; [email protected] Phone (+351) 214 403 500 Applicant – Partner 5 Organisation INRA-MOROCCO Name of contact Afechtal Mohamed, Ph.D.; Bouharroud Rachid, Gender: M (incl. -

The Leafhoppers of Minnesota
Technical Bulletin 155 June 1942 The Leafhoppers of Minnesota Homoptera: Cicadellidae JOHN T. MEDLER Division of Entomology and Economic Zoology University of Minnesota Agricultural Experiment Station The Leafhoppers of Minnesota Homoptera: Cicadellidae JOHN T. MEDLER Division of Entomology and Economic Zoology University of Minnesota Agricultural Experiment Station Accepted for publication June 19, 1942 CONTENTS Page Introduction 3 Acknowledgments 3 Sources of material 4 Systematic treatment 4 Eurymelinae 6 Macropsinae 12 Agalliinae 22 Bythoscopinae 25 Penthimiinae 26 Gyponinae 26 Ledrinae 31 Amblycephalinae 31 Evacanthinae 37 Aphrodinae 38 Dorydiinae 40 Jassinae 43 Athysaninae 43 Balcluthinae 120 Cicadellinae 122 Literature cited 163 Plates 171 Index of plant names 190 Index of leafhopper names 190 2M-6-42 The Leafhoppers of Minnesota John T. Medler INTRODUCTION HIS bulletin attempts to present as accurate and complete a T guide to the leafhoppers of Minnesota as possible within the limits of the material available for study. It is realized that cer- tain groups could not be treated completely because of the lack of available material. Nevertheless, it is hoped that in its present form this treatise will serve as a convenient and useful manual for the systematic and economic worker concerned with the forms of the upper Mississippi Valley. In all cases a reference to the original description of the species and genus is given. Keys are included for the separation of species, genera, and supergeneric groups. In addition to the keys a brief diagnostic description of the important characters of each species is given. Extended descriptions or long lists of references have been omitted since citations to this literature are available from other sources if ac- tually needed (Van Duzee, 1917). -

A Review of the Systematics of Hawaiian Planthoppers (Hemiptera: Fulgoroidea)L
Pacific Science (1997), vol. 51, no. 4: 366-376 © 1997 by University of Hawai'i Press. All rights reserved A Review of the Systematics of Hawaiian Planthoppers (Hemiptera: Fulgoroidea)l MANFRED ASCHE2 ABSTRACT: With 206 endemic species, the phytophagous Fulgoroidea, or planthop pers, are among the most important elements of the native Hawaiian fauna. These principally monophagous or oligophagous insects occur in nearly all Hawaiian terrestrial ecosystems. Species of two of the 18 planthopper families occurring worldwide have successfully colonized and subsequently radiated in Hawai'i. Based on collections made mainly by Perkins, Kirkaldy, Muir, Giffard, and Swezey, more than 95% of these species were described in the first three decades of this century. The systematics of the Hawaiian planthoppers has changed little in the past 60 yr and is not based on any phylogenetic analyses. This paper attempts a preliminary phylogenetic evaluation ofthe native Hawaiian p1anthoppers on the basis ofcompara tive morphology to recognize monophyletic taxa and major evolutionary lines. The following taxa are each descendants of single colonizing species: in Cixiidae, the Hawaiian Oliarus and Iolania species; in De1phacidae, Aloha partim, Dictyophoro delphax, Emoloana, Leialoha + Nesothoe, Nesodryas, and at least four groups within Nesosydne. Polyphyletic taxa are the tribe "Alohini," Aloha s.l., Nesorestias, Nesosydne s.l., and Nothorestias. Non-Hawaiian species currently placed in Iolania, Oliarus, Aloha, Leialoha, and Nesosydne are not closely allied to the Hawaiian taxa. The origin of the Hawaiian planthoppers is obscure. The Hawaiian Oliorus appear to have affinities to (North) American taxa. ALTHOUGH THE HAWAIIAN ISLANDS are the most Other groups of Hawaiian insects have isolated islands on earth, they house a remark received far less attention, although they are ably rich flora and fauna. -
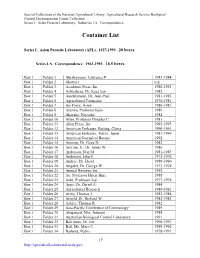
Container List
Special Collections of the National Agricultural Library: Agricultural Research Service Biological Control Documentation Center Collection Series I. Asian Parasite Laboratory. Subseries I.A. Correspondence. Container List Series I. Asian Parasite Laboratory (APL). 1927-1993. 20 boxes. Series I.A. Correspondence. 1963-1993. 16.5 boxes. Box 1 Folder 1 Abrahamson, Lawrence P. 1983-1984 Box 1 Folder 2 Abstract n.d. Box 1 Folder 3 Academic Press, Inc. 1986-1993 Box 1 Folder 4 Achterberg, Dr. Kees van 1982 Box 1 Folder 5 Aeschlimann, Dr. Jean-Paul 1981-1985 Box 1 Folder 6 Agricultural Counselor 1976-1981 Box 1 Folder 7 Air Force, Army 1980-1981 Box 1 Folder 8 Aizawa, Professor Keio 1986 Box 1 Folder 9 Akasaka, Naoyuki 1982 Box 1 Folder 10 Allen, Professor Douglas C. 1981 Box 1 Folder 11 Allen Press, Inc. 1992-1993 Box 1 Folder 12 American Embassy, Beijing, China 1990-1991 Box 1 Folder 13 American Embassy, Tokyo, Japan 1981-1990 Box 1 Folder 14 American Journal of Botany 1992 Box 1 Folder 15 Amman, Dr. Gene D. 1982 Box 1 Folder 16 Amrine, Jr., Dr. James W. 1986 Box 1 Folder 17 Anderson, Don M. 1981-1983 Box 1 Folder 18 Anderson, John F. 1975-1976 Box 1 Folder 19 Andres, Dr. Lloyd 1989-1990 Box 1 Folder 20 Angalet, Dr. George W. 1973-1978 Box 1 Folder 21 Annual Reviews Inc. 1992 Box 1 Folder 22 Ao, Professor Hsien-Bine 1988 Box 1 Folder 23 Aoki, Professor Joji 1977-1978 Box 1 Folder 24 Apps, Dr. Darrel A. 1984 Box 1 Folder 25 Agricultural Research 1980-1981 Box 1 Folder 26 Army, Thomas J. -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000).