Distribution and Geographic Variation in the Western Woolly Lemur (Avahi Occidentalis) with Description of a New Species (A
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fascinating Primates 3/4/13 8:09 AM Ancient Egyptians Used Traits of an Ibis Or a Hamadryas Used Traits Egyptians Ancient ) to Represent Their God Thoth
© Copyright, Princeton University Press. No part of this book may be distributed, posted, or reproduced in any form by digital or mechanical means without prior written permission of the publisher. Fascinating Primates Fascinating The Beginning of an Adventure Ever since the time of the fi rst civilizations, nonhuman primates and people have oc- cupied overlapping habitats, and it is easy to imagine how important these fi rst contacts were for our ancestors’ philosophical refl ections. Long ago, adopting a quasi- scientifi c view, some people accordingly regarded pri- mates as transformed humans. Others, by contrast, respected them as distinct be- ings, seen either as bearers of sacred properties or, conversely, as diabolical creatures. A Rapid Tour around the World In Egypt under the pharaohs, science and religion were still incompletely separated. Priests saw the Papio hamadryas living around them as “brother baboons” guarding their temples. In fact, the Egyptian god Thoth was a complex deity combining qualities of monkeys and those of other wild animal species living in rice paddies next to temples, all able to sound the alarm if thieves were skulking nearby. At fi rst, baboons represented a local god in the Nile delta who guarded sacred sites. The associated cult then spread through middle Egypt. Even- tually, this god was assimilated by the Greeks into Hermes Trismegistus, the deity measuring and interpreting time, the messenger of the gods. One conse- quence of this deifi cation was that many animals were mummifi ed after death to honor them. Ancient Egyptians used traits of an ibis or a Hamadryas Baboon (Papio hamadryas) to represent their god Thoth. -

Avahi Laniger)
A. Zaramody et al.: Phylogeny of Eastern Woolly Lemurs MOLECULAR PHYLOGENY AND TAXONOMIC REVISION OF THE EASTERN WOOLLY LEMURS (AVAHI LANIGER) Zaramody A, Fausser J-L, Roos C, Zinner D, Andriaholinirina N, Rabarivola C, Norscia I, Tattersall I and Rumpler Y Key words: Avahi, Strepsirrhini, taxonomy, mtDNA, cytogenetics, new species Abstract The western and northern populations of woolly lemurs (Avahi) have been di- vided into three distinct species (A. cleesei, A. occidentalis and A. unicolor), whereas the eastern populations are still considered to represent a single species (A. laniger), despite the wider distribution of woolly lemurs in this region. To analyze the diver- sity within the eastern population and among the eastern and western populations, we compared cytogenetic data and mitochondrial DNA (mtDNA) sequences from woolly lemurs from 14 sites in the east of Madagascar and from three sites in the west, representing three of the four recognized species. Cytogenetic and mtDNA data are in agreement and confirm the distinctiveness of A. laniger and A. occiden- talis. Within A. laniger the molecular data revealed large genetic distances among local populations. On the basis of these new data we propose to split A. laniger into three species: (1) north of the Mongoro/Onive Rivers, (2) south of the Mongoro/Onive Rivers at least as far south as Mahasoarivo, and (3) from the south-east (Manombo, Sainte Luce). Within the south-eastern species (3) two clearly separated subspecies can be distinguished, one from the region of Manombo and the other from the region of Sainte Luce. The northern species (1) shows considerable intraspecies genetic dis- tances and may consist of several populations distinguishable as subspecies. -

Lemur Bounce!
Assets – Reections Icon Style CoverAssets Style – Reections 1 Icon Style Y E F O N Cover StyleR O M M R A A Y E F D O N C A S R O GA M M R A D A AGASC LemurVISUAL Bounce! BRAND LearningGuidelines and Sponsorship Pack VISUAL BRAND Guidelines 2 Lemur Bounce nni My name is Lennie Le e Bounce with me B .. ou and raise money to n c protect my Rainforest e L ! home! L MfM L 3 What is a Lemur Bounce? A Lemur Bounce is a sponsored event for kids to raise money by playing bouncing games. In this pack: * Learning fun for kids including facts and quizzes Indoor crafts and outside bouncing games for * the Lemur Bounce Day * Links to teaching resources for schools * Lesson planning ideas for teachers * Everything you need for a packed day of learning and fun! Contents Fun Bounce activities Page No. Let’s BounceBounce – YourLemur valuable support 5 n n Lemur Bouncee i e L – Basics 7 B u o Planningn for a Lemur Bounce Day 8 c L e Lemur! Bounce Day Assembly 8 Make L a Lemur Mask 9 Make a Lemur Tail & Costume 10 Games 11-12 Sponsorship Forms 13-14 Assets – Reections M f Certificates 15-20 Icon Style M Cover Style L Y E F O N R Fun Indoor activities O M M R A Planning your lessons for a Lemur Bounce Day 22 D A A SC GA Fun Facts about Madagascar 23 Fun Facts about Lemurs 24 Colouring Template 25 VISUAL BRAND Guidelines Word Search 26 Know your Lemurs 27 Lemur Quiz 28 Madagascar Quiz 29-31 Resources 32 Song and Dance 33 Contact us 34 LEMUR BOUNCE BOUNCE fM FOR MfM LEMUR BASICS Y NE FO O R WHAT ‘LEM OUNC“ ‘ DO YOU KNOW YOUR LEMUR M Have you ever seen a lemur bounce? Maaasar is ome to over 0 seies o endemi lemurs inluding some M very ouncy ones lie the Siaa lemur ut tese oreous rimates are highl endangere e need to at R Let’s Bounce - Your valuable support A no to sae teir aitat and rotect tem rom etin tion arity Mone or Maaasar is alling out to A D C Wherechildren does everywhere the money to organizego? a fun charity ‘lemur bounce’. -

Coexistence of Confamilial, Folivorous Indriids, Propithecus Diadema And
Washington University in St. Louis Washington University Open Scholarship Arts & Sciences Electronic Theses and Dissertations Arts & Sciences Spring 5-15-2017 Coexistence of Confamilial, Folivorous Indriids, Propithecus diadema and Indri indri, at Betampona Strict Nature Reserve, Madagascar Lana Kerker Oliver Washington University in St. Louis Follow this and additional works at: https://openscholarship.wustl.edu/art_sci_etds Part of the Biodiversity Commons, Biological and Physical Anthropology Commons, Natural Resources and Conservation Commons, and the Natural Resources Management and Policy Commons Recommended Citation Oliver, Lana Kerker, "Coexistence of Confamilial, Folivorous Indriids, Propithecus diadema and Indri indri, at Betampona Strict Nature Reserve, Madagascar" (2017). Arts & Sciences Electronic Theses and Dissertations. 1134. https://openscholarship.wustl.edu/art_sci_etds/1134 This Dissertation is brought to you for free and open access by the Arts & Sciences at Washington University Open Scholarship. It has been accepted for inclusion in Arts & Sciences Electronic Theses and Dissertations by an authorized administrator of Washington University Open Scholarship. For more information, please contact [email protected]. WASHINGTON UNIVERSITY IN ST. LOUIS Department of Anthropology Dissertation Examination Committee Crickette Sanz, Chair Kari Allen Benjamin Z. Freed Jane Phillips-Conroy David Strait Mrinalini Watsa Coexistence of Confamilial, Folivorous Indriids, Propithecus diadema and Indri indri, at Betampona Strict -

Magical Madagascar June 19 – July 1, 2017 Anjajavy Extension July 1 – July 4, 2017 Magical Madagascar June 19 – July 1, 2017
Magical Madagascar June 19 – July 1, 2017 Anjajavy extension July 1 – July 4, 2017 Magical Madagascar June 19 – July 1, 2017 We invite you to be part of this exclusive and remarkable adventure through Madagascar, one of the most extraordinary places on Earth. There is no other trip to Madagascar like this one. In addition to the many splendors of that unique island country, you will experience exclusive areas not open to tourists. The Duke Lemur Center has been working in Madagascar for 30 years and we will have access to rarely visited areas, Duke connected conservation projects and people. You will meet with conservation and research experts, providing a truly unique experience. This trip is perfect for first time Madagascar visitors as well as for those returning to Madagascar for a second or third trip! Madagascar is home to all of the world’s lemurs (some 100+ species) and over half of the world’s chameleons, in addition to more than 170 species of frogs. Every year scientists discover new species of plants and animals in Madagascar’s deserts and forests. There is no question Madagascar is a nature lover's dream and an exceptional photographic destination. The animals we will see in the protected areas are habituated and easily approached, making for terrific photographic opportunities. Our itinerary is designed to cover some of the island’s highlights, including six different eco-systems with a diversity of wildlife and photographic subjects. We have chosen these six stops to showcase the incredible diversity in landscape flora and fauna common to each of these areas. -

Rare Mammals of Madagascar
Rare Mammals of Madagascar Trip Highlights from November 2019 and May 2016 by Ian Loyd Introduction Madagascar must be one of the most extraordinary destinations in the world for those interested in natural history. This record of some stand out sightings and where to find key species will hopefully help those planning a trip. Often referred to as the “Eighth Continent” and “The Big Red Island”, Madagascar is the world’s oldest and fourth largest island and after millions of years of isolation, a wildlife holiday there is truly unlike anywhere else. There is a vast range of ecosystems to explore: wet rainforest, dry tropical deciduous forest and the unique spiny forest found only in Madagascar’s far southwest. In addition, there are also coral reefs and stunning white sand beaches to enjoy along the coast, plus the unique stone forests known as tsingy and fascinating cultures to discover. Nearly all of Madagascar’ staggering biodiversity is found nowhere else and much of it is sadly increasingly threatened with extinction. Wildlife highlights undoubtably include the mysterious nocturnal aye aye, iconic ring-tailed lemur, the beautiful sifaka family, the agile indri, tiny mouse lemurs, charismatic fossa, both giant and miniature chameleons, camouflaged leaf-tailed geckos, colourful frogs and its fantastically varied endemic birds. (2)…/ Wildlife tourism has emerged as a major source of foreign income for Madagascar. By visiting you contribute directly to the conservation of its precious remaining forests with their unique wildlife, and the welfare of its people who are among the poorest (yet friendliest) in the world. Your presence facilitates the hiring of park rangers who guard the precious reserves from illegal logging, slash-and-burn farmers and bush-meat hunters. -

Male-Specific Use of the Purr in the Ring-Tailed Lemur (Lemur Catta)
Original Article Folia Primatol 2014;85:201–214 Received: October 25, 2013 DOI: 10.1159/000363057 Accepted after revision: April 22, 2014 Published online: August 16, 2014 Male-Specific Use of the Purr in the Ring-Tailed Lemur (Lemur catta) Laura M. Bolt Department of Anthropology, University of Toronto, Toronto, Ont. , Canada Key Words Purr · Ring-tailed lemur · Lemur catta · Sex-specific behaviour · Information signal · Male vocalization · Dominance rank · Call rate · Male agonism · Epiphenomenon Abstract In mammals, purring has been described in mostly affiliative contexts. In the ring- tailed lemur (Lemur catta) , both males and females purr, but only males were observed purring in agonistic contexts. In order to determine whether male ring-tailed lemurs purr as aggressive displays during intrasexual agonistic encounters, 480 h of focal data were collected on 25 adult males from Beza Mahafaly Special Reserve, Madagascar, from March to July 2010. The male purring rate increased during periods of male-male ago- nism when compared to times without intrasexual agonism, and the purring rate was positively correlated with male dominance rank. However, the purring rate was not sig- nificantly higher during winning agonistic interactions when compared with losing en- counters. My results indicate that the male ring-tailed lemur purr is used most frequent- ly as an agonistic vocalization in male-male encounters, in addition to being used less frequently in other social contexts, including during tail-waving at females, resting, scent-marking, feeding and copulation. Dominant males have higher purring rates across social situations, suggesting that the purring rate may be driven by intrinsic male qualities rather than functioning as a meaningful signal in each disparate social context. -

Lemurs of Madagascar – a Strategy for Their
Cover photo: Diademed sifaka (Propithecus diadema), Critically Endangered. (Photo: Russell A. Mittermeier) Back cover photo: Indri (Indri indri), Critically Endangered. (Photo: Russell A. Mittermeier) Lemurs of Madagascar A Strategy for Their Conservation 2013–2016 Edited by Christoph Schwitzer, Russell A. Mittermeier, Nicola Davies, Steig Johnson, Jonah Ratsimbazafy, Josia Razafindramanana, Edward E. Louis Jr., and Serge Rajaobelina Illustrations and layout by Stephen D. Nash IUCN SSC Primate Specialist Group Bristol Conservation and Science Foundation Conservation International This publication was supported by the Conservation International/Margot Marsh Biodiversity Foundation Primate Action Fund, the Bristol, Clifton and West of England Zoological Society, Houston Zoo, the Institute for the Conservation of Tropical Environments, and Primate Conservation, Inc. Published by: IUCN SSC Primate Specialist Group, Bristol Conservation and Science Foundation, and Conservation International Copyright: © 2013 IUCN Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the source is fully acknowledged. Reproduction of this publication for resale or other commercial purposes is prohibited without prior written permission of the copyright holder. Inquiries to the publisher should be directed to the following address: Russell A. Mittermeier, Chair, IUCN SSC Primate Specialist Group, Conservation International, 2011 Crystal Drive, Suite 500, Arlington, VA 22202, USA Citation: Schwitzer C, Mittermeier RA, Davies N, Johnson S, Ratsimbazafy J, Razafindramanana J, Louis Jr. EE, Rajaobelina S (eds). 2013. Lemurs of Madagascar: A Strategy for Their Conservation 2013–2016. Bristol, UK: IUCN SSC Primate Specialist Group, Bristol Conservation and Science Foundation, and Conservation International. 185 pp. ISBN: 978-1-934151-62-4 Illustrations: © Stephen D. -

Madagascar's Lemurs
Madagascar's Lemurs Naturetrek Tour Report 10 - 23 November 2012 Black and White Ruffed Lemur Hubbard's Sportive Lemur Spiny Backed Chameleon Red-bellied Lemur Report compiled by Nick Acheson Images by Kerrie Warburton Naturetrek Cheriton Mill Cheriton Alresford Hampshire SO24 0NG England T: +44 (0)1962 733051 F: +44 (0)1962 736426 E: [email protected] W: www.naturetrek.co.uk Tour Report Madagascar's Lemurs Tour Leaders: Claude Rambeloson Kerrie Warburton Nick Acheson Participants: Mary Ashwin Alison Higgins Pauline Davis Sue Levings Sonia Hubbard Frosso Miltiadou Chris Evans Helen Pinchin Julie Ives Ann Mellor Peter Phillips Julianne Schwenke Day 0 Saturday 10th November Leave the UK Day 1 Sunday 11th November What better way to begin a Madagascar’s Lemurs tour than by getting to know the many lemurs at Lemurs Park. These captive animals have all been rescued from unsuitable conditions as pets and now live in an extensive patch of pine and eucalyptus trees which is slowly being converted to native vegetation. The lemurs here are healthy and relaxed and have the photographic advantage of being dippily tame. So, shamelessly, we cooed over Mongoose and Common Brown Lemurs, we giggled over the antics of families of Coquerel’s Sifakas, and we swooned at the sheer handsomeness of the pair of Black-and-White Ruffed Lemurs which had recently had a happy event. There was plenty of wild wildlife here too. A brilliant make Souimanga Sunbird shone as he sang in the sunshine, young Madagascar Kestrels poked their heads from the nest (in an old Hamerkop nest) as their parents came and went, and both Ornate and Broad-tailed Plated Lizards trotted along the paths in front of us. -
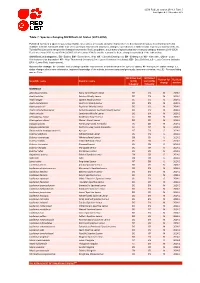
Table 7: Species Changing IUCN Red List Status (2013-2014)
IUCN Red List version 2014.3: Table 7 Last Updated: 13 November 2014 Table 7: Species changing IUCN Red List Status (2013-2014) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2013 (IUCN Red List version 2013.2) and 2014 (IUCN Red List version 2014.3) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.); E - Previous listing was an Error. IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2013) List (2014) change version Category Category MAMMALS Allocebus trichotis Hairy-eared Dwarf Lemur DD VU N 2014.1 Avahi betsileo Betsileo Woolly Lemur -

Lemurs of Madagascar and the Comoros the Lucn Red Data Book
Lemurs of Madagascar and the Comoros The lUCN Red Data Book Prepared by The World Conservation Monitoring Centre lUCN - The World Conservation Union 6(^2_ LEMURS OF MADAGASCAR and the Comoros The lUCN Red Data Book lUCN - THE WORLD CONSERVATION UNION Founded in 1948, lUCN - The World Conservation Union - is a membership organisation comprising governments, non-governmental organisations (NGOs), research institutions, and conservation agencies in 120 countries. The Union's objective is to promote and encourage the protection and sustainable utilisation of living resources. Several thousand scientists and experts from all continents form part of a network supporting the work of its six Commissions: threatened species, protected areas, ecology, sustainable development, environmental law and environmental education and training. Its thematic programmes include tropical forests, wetlands, marine ecosystems, plants, the Sahel, Antarctica, population and natural resources, and women in conservation. These activities enable lUCN and its members to develop sound policies and programmes for the conservation of biological diversity and sustainable development of natural resources. WCMC - THE WORLD CONSERVATION MONITORING CENTRE The World Conservation Monitoring Centre (WCMC) is a joint venture between the three partners in the World Conservation Strategy, the World Conservation Union (lUCN), the World Wide Fund for Nature (WWF), and the United Nations Environment Programme (UNEP). Its mission is to support conservation and sustainable development by collecting and analysing global conservation data so that decisions affecting biological resources are based on the best available information. WCMC has developed a global overview database of the world's biological diversity that includes threatened plant and animal species, habitats of conservation concern, critical sites, protected areas of the world, and the utilisation and trade in wildlife species and products. -

LEMUR NEWS the Newsletter of the Madagascar Section of the I.U.C.N./S.S.C
LEMUR NEWS The Newsletter of the Madagascar Section of the I.U.C.N./S.S.C. Primate Specialist Group NUMBER 12, June 2007 DPZ Deutsches Primatenzentrum Universität Hamburg Cover photo: Microcebus sp. from Andasibe (photo by Ute Meede). LEMUR NEWS The Newsletter of the Madagascar Section of the IUCN/SSC Primate Specialist Group PSG Chairman: Russel A. Mittermeier PSG Deputy Chairman: Anthony B. Rylands Addresses for contributions Jörg U. Ganzhorn Editors Abt. Tierökologie und Naturschutz Jörg U. Ganzhorn Universität Hamburgl Ken Glander Martin-Luther-King-Platz 3 Jonah Ratsimbazafy 20146 Hamburg Rodin M. Rasoloarison Germany Michael Schwibbe E-mail: [email protected] Anne Yoder Jonah Ratsimbazafy, Rodin M. Rasoloarison, Assistant Editors Rose Marie Randrianarison Anja Ganzhorn GERP Rose Marie Randrianarison 34, Cité des Professeurs, Fort Duchesne Antananarivo (101) Layout Madagascar Heike Klensang E-mail: [email protected] Lemur News online Number of copies: 1200 Volumes 3 -present are available online at www.dpz.eu ISSN 1608-1439 Printed by Goltze GmbH & Co. KG, Göttingen, Germany Lemur News Vol. 12, 2007 Page 1 EDITORIAL ERRATUM The last few months have brought exciting news. Mada- There is a mistake in the legend of Figure 1 in the article gascar’s old and new president, Marc Ravalomanana, by Rabarivola et al., Lemur News 11: page 46: we apolo- has made remarkable statements with respect to hu- gize to the authors and to the readers for not having man population growth, new regulations for hunting paid better attention. have been established, many new forms of lemurs have been described, and last, but not least the regulations for doctoral candidates within the Malagasy university system have changed.