Advances in Engineering of Fluorescent Proteins And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Monitoring of Switches in Heterochromatin-Induced Silencing Shows Incomplete Establishment and Developmental Instabilities
Monitoring of switches in heterochromatin-induced silencing shows incomplete establishment and developmental instabilities Farah Bughioa, Gary R. Huckellb,1, and Keith A. Maggerta,2 aDepartment of Cellular and Molecular Medicine, University of Arizona, Tucson, AZ 85724; and bPrivate address, San Diego, CA 92120 Edited by James A. Birchler, University of Missouri, Columbia, MO, and approved August 28, 2019 (received for review June 6, 2019) Position effect variegation (PEV) in Drosophila results from new While the natural factors that influence establishment have juxtapositions of euchromatic and heterochromatic chromosomal only recently begun to be explored (34, 35), multiple models for regions, and manifests as striking bimodal patterns of gene ex- maintenance have been proposed, including DNA cytosine pression. The semirandom patterns of PEV, reflecting clonal rela- methylation and histone modifications stably inherited through tionships between cells, have been interpreted as gene-expression S-phase (36–39). The former does not exist in the common yeast states that are set in development and thereafter maintained model systems, in Caenorhabditis elegans or in Drosophila (40, 41; without change through subsequent cell divisions. The rate of in- cf. refs. 42 and 43), and so it is widely accepted that epigenetic stability of PEV is almost entirely unexplored beyond the final gene-regulatory (e.g., heterochromatin-induced silencing) infor- expression of the modified gene; thus the origin of the expressiv- mation must be encoded by the histone modifications in these ity and patterns of PEV remain unexplained. Many properties of model systems. Histone modifications (and DNA methylation) PEV are not predicted from currently accepted biochemical and are part of the mechanisms of activation or repression, although theoretical models. -

KDM7 Is a Dual Demethylase for Histone H3 Lys 9 and Lys 27 and Functions in Brain Development
Downloaded from genesdev.cshlp.org on October 1, 2021 - Published by Cold Spring Harbor Laboratory Press RESEARCH COMMUNICATION elegans to humans and is characterized by the presence of KDM7 is a dual demethylase a PHD-type zinc finger motif in addition to the JmjC do- for histone H3 Lys 9 and Lys main (Supplemental Fig. S1A). Whereas the human genes for PHF8 and PHF2 are associated with X-linked mental 27 and functions in brain retardation and hereditary sensory neuropathy type I, respectively (Hasenpusch-Theil et al. 1999; Laumonnier development et al. 2005; Abidi et al. 2007; Koivisto et al. 2007), little is Yu-ichi Tsukada,1,2,3 Tohru Ishitani,4 known about KIAA1718. Bioinformatic analysis of the 1,2,5 JmjC domains of KIAA1718, PHF8, and PHF2 indicated and Keiichi I. Nakayama that predicted Fe(II)- and a-ketoglutarate (a-KG)-binding 1Division of Cell Biology, Medical Institute of Bioregulation, sites are conserved, with the exception of the former in Kyushu University, Higashi-ku, Fukuoka 812-8582, Japan; PHF2, and that they share extensive similarity with the 2CREST, Japan Science and Technology Agency (JST), JmjC domain of JHDM1/KDM2 (Supplemental Fig. S1B). Kawaguchi, Saitama 332-0012, Japan; 3PRESTO, Japan Science Conservation of residues within the putative cofactor- and Technology Agency (JST), Kawaguchi, Saitama 332-0012, binding sites of KIAA1718 suggested that this protein Japan; 4Division of Cell Regulation Systems, Medical Institute might possess histone demethylase activity, and there- of Bioregulation, Kyushu University, Higashi-ku, Fukuoka fore might also contribute to transcriptional regulation of 812-8582, Japan genes in the nervous system. -

Single-Molecule Spectroscopy and Imaging of Biomolecules in Living Cells
Anal. Chem. 2010, 82, 2192–2203 Single-Molecule Spectroscopy and Imaging of Biomolecules in Living Cells Samuel J. Lord, Hsiao-lu D. Lee, and W. E. Moerner* Department of Chemistry, Stanford University, Stanford, California 94305-5080 The number of reports per year on single-molecule imaging instance, the shape of the distribution may be skewed or reveal experiments has grown roughly exponentially since the first multiple subpopulations, which may offer insight into underlying successful efforts to optically detect a single molecule were mechanisms. Each single molecule is a local reporter on the completed over two decades ago. Single-molecule spectros- makeup and conditions of its immediate surroundings, its “na- copy has developed into a field that includes a wealth of noenvironment”, and thus acts as a readout of spatial heterogene- experiments at room temperature and inside living cells. The ity of a sample. SMS also measures time-dependent processes that fast growth of single-molecule biophysics has resulted from are not necessarily synchronized throughout the sample or its benefits in probing heterogeneous populations, one population. For example, multiple catalytic states of an enzyme molecule at a time, as well as from advances in microscopes will be convolved with all the states of other copies in an ensemble, and detectors. This Perspective summarizes the field of live- whereas a SMS experiment can measure uncorrelated stochastic cell imaging of single biomolecules. transitions of a single enzyme. SMS also has the ability to observe intermediate states or rare events, given that the instruments have Single-molecule biophysics spans a range of experiments, from sufficient time resolution. -

Rational Design of Small Molecule Fluorescent Probes for Biological Applications
Organic & Biomolecular Chemistry Rational Design of Small Molecule Fluorescent Probes for Biological Applications Journal: Organic & Biomolecular Chemistry Manuscript ID OB-REV-06-2020-001131.R1 Article Type: Review Article Date Submitted by the 13-Jul-2020 Author: Complete List of Authors: Jun, Joomyung; University of Pennsylvania, Chemistry; Massachusetts Institute of Technology, Chemistry Chenoweth, David; University of Pennsylvania, Department of Chemistry Petersson, E.; University of Pennsylvania, Chemistry Page 1 of 16 Organic & Biomolecular Chemistry ARTICLE Rational Design of Small Molecule Fluorescent Probes for Biological Applications a,b a a,c Received 00th January 20xx, Joomyung V. Jun, David M. Chenoweth* and E. James Petersson* Accepted 00th January 20xx Fluorescent small molecules are powerful tools for visualizing biological events, embodying an essential facet of chemical DOI: 10.1039/x0xx00000x biology. Since the discovery of the first organic fluorophore, quinine, in 1845, both synthetic and theoretical efforts have endeavored to “modulate” fluorescent compounds. An advantage of synthetic dyes is the ability to employ modern organic chemistry strategies to tailor chemical structures and thereby rationally tune photophysical properties and functionality of the fluorophore. This review explores general factors affecting fluorophore excitation and emission spectra, molar absorption, Stokes shift, and quantum efficiency; and provides guidelines for chemist to create novel probes. Structure- property relationships concerning the substituents are discussed in detail with examples for several dye families. Then, we present a survey of functional probes based on PeT, FRET, and environmental or photo-sensitivity, focusing on representative recent work in each category. We believe that a full understanding of dyes with diverse chemical moieties enables the rational design of probes for the precise interrogation of biochemical and biological phenomena. -

Product Information Sheet
AAT Bioquest®, Inc. Product Technical Information Sheet Last Updated July 2012 Classic Calcium Detection Reagents Calcium acts as a universal second messenger in a variety of cells. Numerous functions of all types of cells are regulated by Ca2+ to a greater or lesser degree, thus calcium measurement is critical for numerous biological investigations. Since the 1920s, scientists have attempted to measure Ca2+, but few were successful due to limited availability of Ca2+ probes. The first reliable measurement of Ca2+ was performed by Ridgway and Ashley by injecting the photoprotein aequorin into the giant muscle fiber of the barnacle. Subsequently, in the 1980s, Tsien and colleagues produced a variety of fluorescent indicators. Among them Indo-1, Fura-2, Fluo-3 and Rhod-2 have been the most valuable dyes for measuring Ca2+ with a fluorescence instrument. Fluorescent probes that show spectral responses upon binding to Ca2+ have enabled researchers to investigate changes in intracellular free Ca2+ concentrations by using fluorescence microscopy, flow cytometry, fluorescence spectroscopy and fluorescence microplate readers. Most of these fluorescent indicators are derivatives of BAPTA chelators that incorporate a PET system responsive to calcium. FLIPR® and FlexStation™ instruments of Molecular Devices Corp., FDSS of Hamamatsu Corp. and NOVOstar™ of BMG Technologies have enabled high throughput measurement of calcium for GPCR and ion channel research. There are quite a few factors that need to be considered when selecting a fluorescent Ca2+ indicator. Spectral Properties: For UV excitation, Indo-1 and Fura-2 are widely used. Fluo-3 is preferred for 488 nm excitation while Rhod-2 and X-rhod are used for red emissions. -

GFP and GFP-Like Proteins
REVIEW ARTICLE Evolving trends in biosciences: multi-purpose proteins – GFP and GFP-like proteins Mythili Krishna1 and Baban Ingole2,* 1Institute of Science and Technology, JNT University, Kukatpally, Hyderabad 500 072, India 2National Institute of Oceanography, Dona Paula, Goa 403 004, India (Figure 2). This pioneering work was conducted by Shimo- The sea is considered as holding a clue to many known and unknown biologically active compounds. A family mura and co-workers in the early 1960s. Later, in 1971 of protein named Green Fluorescent Proteins (GFP)- like proteins, initially isolated from marine organisms, started a trend in biotechnological research, which is expanding day-by-day. A gross review of the same is presented in this article dealing with their occurrence, chemistry, applications, phylogenic analysis and the Indian perspective. These proteins are present in a wide variety of marine organisms, from corals to jelly- fish. Chromophore in these proteins is composed of three amino acid residues, Ser65–Try66–Gly67, and requires molecular oxygen for its maturation. Its applications include use in in vivo imaging as well as in plant biology. Thus, the protein has become one of the most important tools used in contemporary biosci- ences. Though the very first protein identified was from the jellyfish Aequorea victoria in the 1960s, the list is ever-expanding with not only the fluorescent but also the non-fluorescent proteins being connected to the same superfamily. Hence, they also offer help in the phylogenetic analysis of different organisms, to know which period of evolution has diverted a parti- Figure 1. Jellyfish, Aequorea victoria. [Figure reprinted from ref. -

Recent Advances in Chemical Biology Using Benzophenones and Diazirines As Radical Precursors
molecules Review Recent Advances in Chemical Biology Using Benzophenones and Diazirines as Radical Precursors 1,2, , 1,2, Muhammad Murtaza Hassan * y and Olasunkanmi O. Olaoye y 1 Department of Chemical and Physical Sciences, University of Toronto Mississauga, 3359 Mississauga Road North, Mississauga, ON L5L 1C6, Canada; [email protected] 2 Department of Chemistry, University of Toronto, 80 St. George Street, Toronto, ON M5S 3H6, Canada * Correspondence: [email protected]; Tel.: +1-905-569-4588 These authors contributed equally to this work. y Academic Editor: Edward Lee-Ruff Received: 18 April 2020; Accepted: 9 May 2020; Published: 13 May 2020 Abstract: The use of light-activated chemical probes to study biological interactions was first discovered in the 1960s, and has since found many applications in studying diseases and gaining deeper insight into various cellular mechanisms involving protein–protein, protein–nucleic acid, protein–ligand (drug, probe), and protein–co-factor interactions, among others. This technique, often referred to as photoaffinity labelling, uses radical precursors that react almost instantaneously to yield spatial and temporal information about the nature of the interaction and the interacting partner(s). This review focuses on the recent advances in chemical biology in the use of benzophenones and diazirines, two of the most commonly known light-activatable radical precursors, with a focus on the last three years, and is intended to provide a solid understanding of their chemical and biological principles and their applications. Keywords: photoaffinity labelling; benzophenone; diazirine; radical precursors; interactome; SABRE; hyperpolarizing agents; crosslinking; photochemistry 1. Introduction The use of radicals or photoactivatable radical precursors has become ubiquitous in the fields of medicinal chemistry and chemical biology in the past three decades. -

Ca2+-Activated Kca3.1 Potassium Channels Contribute to the Slow
www.nature.com/scientificreports OPEN Ca2+‑activated KCa3.1 potassium channels contribute to the slow afterhyperpolarization in L5 neocortical pyramidal neurons M. V. Roshchin, V. N. Ierusalimsky, P. M. Balaban & E. S. Nikitin* Layer 5 neocortical pyramidal neurons are known to display slow Ca2+‑dependent afterhyperpolarization (sAHP) after bursts of spikes, which is similar to the sAHP in CA1 hippocampal cells. However, the mechanisms of sAHP in the neocortex remain poorly understood. Here, we identifed the Ca2+‑gated potassium KCa3.1 channels as contributors to sAHP in ER81‑positive neocortical pyramidal neurons. Moreover, our experiments strongly suggest that the relationship between sAHP and KCa3.1 channels in a feedback mechanism underlies the adaptation of the spiking frequency of layer 5 pyramidal neurons. We demonstrated the relationship between KCa3.1 channels and sAHP using several parallel methods: electrophysiology, pharmacology, immunohistochemistry, and photoactivatable probes. Our experiments demonstrated that ER81 immunofuorescence in layer 5 co‑localized with KCa3.1 immunofuorescence in the soma. Targeted Ca2+ uncaging confrmed two major features of KCa3.1 channels: preferential somatodendritic localization and Ca2+‑driven gating. In addition, both the sAHP and the slow Ca2+‑induced hyperpolarizing current were sensitive to TRAM‑ 34, a selective blocker of KCa3.1 channels. Te neocortex is the largest part of the cortex (~ 90% of the entire cortex in humans), and plays a crucial role in higher functions of the brain such as interpretation of sensory information, formation and storage of long-term memory, language usage, and control of voluntary movements. Pyramidal neurons play a key role in all cortical activities and constitute ~ 80% of all neocortical neurons. -

A Coordinated Local Translational Control Point at the Synapse Involving Relief from Silencing and MOV10 Degradation
Neuron Article A Coordinated Local Translational Control Point at the Synapse Involving Relief from Silencing and MOV10 Degradation Sourav Banerjee,1 Pierre Neveu,1,2 and Kenneth S. Kosik1,* 1Neuroscience Research Institute and Department of Cellular Molecular and Developmental Biology 2Kavli Institute for Theoretical Physics University of California, Santa Barbara, Santa Barbara, CA 93106, USA *Correspondence: [email protected] DOI 10.1016/j.neuron.2009.11.023 SUMMARY 2003; Yi and Ehlers, 2005). These observations suggest a link between localized protein degradation and synthesis in the Persistent changes in synaptic strength are locally regulation of synaptic plasticity. In support of this link, protea- regulated by both protein degradation and synthesis; some blockade can diminish late-phase LTP (L-LTP) induced however, the coordination of these opposing limbs at Schaffer collateral-CA1 synapses, like pharmacological inhibi- is poorly understood. Here, we found that the RISC tion of translation (Fonseca et al., 2006). Interestingly, coapplica- protein MOV10 was present at synapses and was tion of proteasome blockers with translation inhibitors largely rapidly degraded by the proteasome in an NMDA- restores L-LTP, suggesting the need for a crucial balance between protein degradation and synthesis for maintenance of receptor-mediated activity-dependent manner. We L-LTP (Fonseca et al., 2006). Furthermore, activity-dependent designed a translational trap to capture those mRNAs relocation of the proteasome from the dendritic shaft to the whose spatiotemporal translation is regulated by synaptic spine resulted in a spatially precise loss of a degradation MOV10. When MOV10 was suppressed, a set of reporter (Bingol and Schuman, 2006). The relocation of UPS mRNAs—including a-CaMKII, Limk1, and the depal- components is reminiscent of polyribosome redistribution in mitoylating enzyme lysophospholipase1 (Lypla1)— potentiated spines (Ostroff et al., 2002) and suggests local selectively entered the polysome compartment. -
Calcium Detection Probes & Assay Kits
Calcium Detection Probes & Assay Kits 2016-2017 Cal-520™ Cal-590™ Fluo-8® AAT Bioquest® Advancing Assay & Test Technologies Our Mission AAT Bioquest® is committed to constantly meet or exceed its customer’s requirements by providing consistently high quality products and services, and by encouraging continuous improvements in its long-term and daily operations. Our core value is Innovation and Customer Satisfaction. Our Story AAT Bioquest®, Inc. (formerly ABD Bioquest, Inc.) develops, manufactures and markets bioanalytical research reagents and kits to life sciences research, diagnostic R&D and drug discovery. We specialize in photometric detections including absorption (color), fluorescence and luminescence technologies. The Company's superior products enable life science researchers to better under- stand biochemistry, immunology, cell biology and molecular biology. AAT Bioquest offers a rapidly expanding list of enabling products. Besides the standard catalog products, we also offer custom services to meet the distinct needs of each customer. Our current services include custom synthesis of biological detection probes, custom development of biochemical, cell-based and diagnostic assays and custom high throughput screening of drug discovery targets. It is my greatest pleasure to welcome you to AAT Bioquest. We greatly appreciate the constant support of our valuable customers. While we continue to rapidly expand, our core value remains the same: Innovation and Customer Satisfaction. We are committed to being the leading provider of novel biological detection solutions. We promise to extend these values to you during the course of our service and to continue to support you with our new products and services. It is our greatest honor to receive valuable feedbacks and suggestions from you so that we can better serve your projects. -

Optical Highlighter Fluorescent Proteins
Contact Us | Carl Zeiss Education in Microscopy and Digital Imaging ZEISS Home ¦ Products ¦ Solutions ¦ Support ¦ Online Shop ¦ ZEISS International ZEISS Campus Home Interactive Tutorials Basic Microscopy Spectral Imaging Spinning Disk Microscopy Optical Sectioning Superresolution Introduction Article Quick Links Introduction Live-Cell Imaging The photophysical properties of fluorescent proteins are often extremely Photoactivation Fluorescent Proteins complex and can involve several distinct emissive and non-emissive (dark) Photoconversion states, as well as on-and-off blinking behavior when observed at the single Microscope Light Sources Photoswitching molecule level. In fact, one of the first observations of photoswitching in Digital Image Galleries fluorescent proteins was reported in wild-type GFP and several of its FP Timers Applications Library enhanced derivatives. As we will see, the photoswitching properties can Print Version allow the investigator to change the color or the emission state of a Reference Library fluorescent protein, providing unique opportunities to track the dynamic behavior of proteins in living cells and animals. This quality can be extremely useful since, unlike the standard fluorescent proteins that are uniformly fluorescent from the time they are produced, Search photosensitive fluorescent proteins can be switched on at a particular time and location within the cell to track the behavior of a tagged protein. Although fluorescent proteins are known to undergo a variety of light-induced switching characteristics, the most useful are photoactivation, photoconversion, and photoswitching, functions that are collectively termed optical highlighting. Photoactivatable fluorescent proteins are capable of being activated from very low level to bright fluorescence emission upon illumination with ultraviolet or violet light, whereas photoconvertable fluorescent proteins can be optically converted from one fluorescence emission bandwidth to another. -
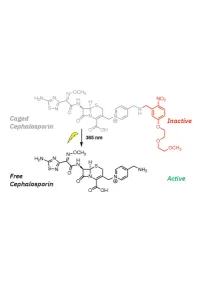
1 20 Introduction
1 Design, Synthesis, and Biological Evaluation of Light-Activated Antibiotics 2 3 Inga S. Shchelik, Andrea Tomio, and Karl Gademann 4 Department of Chemistry, University of Zurich, Winterthurerstrasse 190, 8057, Zurich, SwitzerlanD 5 6 7 ABSTRACT 8 The spatial anD temporal control of bioactivity of small molecules by light (photopharmacology) constitutes a 9 promising approach for stuDy of biological processes and ultimately for the treatment of Diseases. In this stuDy, we 10 investigateD two Different ‘cageD’ antibiotic classes that can unDergo remote activation with UV-light at λ=365 nm, 11 via the conjugation of deactivating and photocleavable units through a short synthetic sequence. The two wiDely useD 12 antibiotics vancomycin and cephalosporin were thus enhanceD in their performance by renDering them 13 photoresponsive and thus suppressing undesired off-site activity. The antimicrobial activity against Bacillus subtilis 14 ATCC 6633, Staphylococcus aureus ATCC 29213, S. aureus ATCC 43300 (MRSA), Escherichia coli ATCC 25922, 15 and Pseudomonas aeruginosa ATCC 27853 could be spatiotemporally controlleD with light. Both molecular series 16 displayed a good activity winDow. The vancomycin Derivative DisplayeD excellent values against Gram-positive 17 strains after uncaging, and the next-generation caged cephalosporin derivative achieved good and broad activity 18 against both Gram-positive and Gram-negative strains after photorelease. 19 Key worDs: antibacterial agents, photopharmacology, photocaging, vancomycin, cephalosporin. 1 20