Genome Profiling of SARS-Cov-2 in Indonesia, ASEAN and the Neighbouring East Asian Countries
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

World Higher Education Database Whed Iau Unesco
WORLD HIGHER EDUCATION DATABASE WHED IAU UNESCO Página 1 de 438 WORLD HIGHER EDUCATION DATABASE WHED IAU UNESCO Education Worldwide // Published by UNESCO "UNION NACIONAL DE EDUCACION SUPERIOR CONTINUA ORGANIZADA" "NATIONAL UNION OF CONTINUOUS ORGANIZED HIGHER EDUCATION" IAU International Alliance of Universities // International Handbook of Universities © UNESCO UNION NACIONAL DE EDUCACION SUPERIOR CONTINUA ORGANIZADA 2017 www.unesco.vg No paragraph of this publication may be reproduced, copied or transmitted without written permission. While every care has been taken in compiling the information contained in this publication, neither the publishers nor the editor can accept any responsibility for any errors or omissions therein. Edited by the UNESCO Information Centre on Higher Education, International Alliance of Universities Division [email protected] Director: Prof. Daniel Odin (Ph.D.) Manager, Reference Publications: Jeremié Anotoine 90 Main Street, P.O. Box 3099 Road Town, Tortola // British Virgin Islands Published 2017 by UNESCO CENTRE and Companies and representatives throughout the world. Contains the names of all Universities and University level institutions, as provided to IAU (International Alliance of Universities Division [email protected] ) by National authorities and competent bodies from 196 countries around the world. The list contains over 18.000 University level institutions from 196 countries and territories. Página 2 de 438 WORLD HIGHER EDUCATION DATABASE WHED IAU UNESCO World Higher Education Database Division [email protected] -

Kajian Interaksi Obat Antihipertensi Pada Pasien Hemodialisis Di Rumah Sakit Umum Yarsi Pontianak Tahun 2017
KAJIAN INTERAKSI OBAT ANTIHIPERTENSI PADA PASIEN HEMODIALISIS DI RUMAH SAKIT UMUM YARSI PONTIANAK TAHUN 2017 Salfitri*1, Nurmainah1, Muhammad Akib Yuswar1 1Program Studi Farmasi Fakultas Kedokteran Universitas Tanjungpura Pontianak Jl Prof. Dr. H. Hadari Nawawi, Kota Pontianak, Kalimantan Barat. ABSTRAK Penyakit gagal ginjal kronik (GGK) merupakan penyakit ginjal stadium akhir. Pasien GGK memerlukan terapi pengganti ginjal seperti hemodialisis. Selain hemodialisis, pasien GGK juga diberikan terapi obat secara polifarmasi yang terdiri dari obat antihipertensi dan golongan obat lainnya. Penelitian ini bertujuan untuk mendeskripsikan jenis obat antihipertensi yang sering berinteraksi dan mengkaji interaksi obat berdasarkan mekanisme kerja serta tingkat keparahan pada pasien GGK yang menjalani hemodialisis. Penelitian ini merupakan penelitian observasional dengan rancangan studi potong lintang (cross sectional) yang bersifat deskriptif, pengumpulan data dilakukan secara retrospektif berdasarkan catatan rekam medik pasien. Data peresepan yang didapat pasien GGK sebelum menjalani hemodialisis di analisis menggunakan software drugs.com. Hasil penelitian menunjukkan bahwa dari 32 sampel terdapat kejadian interaksi obat sebesar 37,50%. Obat yang paling sering berinteraksi adalah golongan CCB yaitu amlodipin sebesar 45,83% dan golongan ACEI yaitu kaptopril sebesar 33,33%. Interaksi obat yang terjadi berdasarkan mekanisme kerja obat yaitu farmakodinamik sebesar 79,17%, farmakokinetik sebesar 8,33% dan yang tidak diketahui mekanisme interaksinya sebesar 12,50%. Sedangkan berdasarkan tingkat keparahannya yaitu minor sebesar 29,17%, moderat sebesar 62,50% dan mayor sebesar 8,33%. Kesimpulan dari penelitian ini adalah obat antihipertensi yang paling banyak digunakan yaitu CCB, sedangkan interaksi obat yang sering terjadi yaitu mekanisme kerja farmakodinamik dan tingkat keparahannya moderat. Kata kunci: Gagal ginjal kronik, Hemodialisis, Interaksi obat. Penulis : Salfitri Program Studi Farmasi Fakultas Kedokteran Universitas Tanjungpura Pontianak Jl. -

International Newsletter
No. 25 October 2010 = Foreword = International Research Activities at RISH in 2010 Professor Hiroshi Yamakawa Chair of the International Academic Exchange Committee, RISH, Kyoto University Humanospheric science is a new between Professor H. A. Muin, Dean of the was held at Gadjah Mada University Club, interdisciplinary science, the goal of Faculty of Forestry, Tanjungpura Universi- Yogyakarta, Indonesia, on June 10-12, which is to conduct research concerning ty, Pontianak, West Kalimantan, Indonesia, 2010. The purpose of the HSS2010 was to a “humanosphere” comprising of “four and Professor T. Tsuda, Director of RISH. share the state-of-the-art research results spheres” including outer space, the atmo- Tanjungpura University is an Indonesian in humanosphere sciences among Indone- sphere, the “forest-sphere”, and the human national university locating in Pontianak, sian and Japanese young scientists. With living environment, all of which are vital the capital city of West Kalimantan Prov- a total of 116 participants from Indonesia to human existence. The Research Institute ince. The research collaboration between (111) and Japan (5), the HSS2010 was for Sustainable Humanosphere (RISH) RISH and the Faculty of Forestry, Tanjung- very successful and inspiring. The research of Kyoto University was established in pura University, was started in 1996 under introductions from the five RISH students, 2004 to promote this new science with the JSPS-LIPI Core University Program in Ms. M. Yokozeki, Mr. S. Nonaka, Ms. M. academic activities and education through the field of wood science, and that program Nishioka, Mr. R. S. Maail, and Mr. Y. P. domestic and international collaborative ran from 1996 to 2005. -

Improving Bahasa Competence Through Fast Track-Based Participatory Model
Improving Bahasa Competence through Fast Track-Based Participatory Model Try Hariadi1,2, Sarwiji Suwandi1, St. Y. Slamet1 and Sumarwati Sumarwati1 1Universitas Sebelas Maret, Surakarta, Indonesia 2IKIP PGRI Pontianak, Pontianak, Indonesia [email protected] Keywords: Linguistic competence, Bahasa Indonesia, participatory model, fast track, action research. Abstract: The problem found in higher education especially in learning Bahasa is that the undergraduates cannot meet the expected language learning outcome. This research aims to improve Bahasa competence through fast track-based participatory model in Tanjungpura University, Indonesia. The subjects of the research are the lecturer of Bahasa Education Department and the undergraduates of Chemistry Education Department, teacher training and education faculty, Tanjungpura University in the academic year of 2017/2018. This three- cycle classroom action involves some steps as planning, acting, observing, and reflecting. The instruments of the research are Bahasa competence test, field note, and observation guideline. The data are analyzed quantitatively and qualitatively. The research findings show that after joining the lectures: (1) there is improvement on undergraduates’ linguistic competence; almost 90% undergraduates get score >70 and (2) more than 70% undergraduates pass the test. The undergraduates’ Bahasa competence is improved since there are more of them that participate through various tracks in listening, speaking, reading, and writing. Implementation of participatory model based on fast track gives positive impact with the improvement of Indonesian language competence of students in the learning of general basic subjects of Indonesian language. 1 INTRODUCTION al., 2013). Fast track done early can improve the expected learning outcome and lessen the gap. It Bahasa competence is a type of competence that focuses on individual learning that is suitable with the covers listening, speaking, reading, and writing skill learning purpose (Turrent, 2009). -

8Th Batch of SEA-Teacher Student Exchange: Good Practices and Lesson Learned Chiang Rai Rajabhat University, Thailand (CRRU) Background
8th Batch of SEA-Teacher Student Exchange: Good Practices and Lesson Learned Chiang Rai Rajabhat University, Thailand (CRRU) Background CRRUjoined Sea Teacher Project since Bat ch 2 (2016) and consistently participated in this program up tothe current batch (Bat ch 8, 2019). CRRU Outbound-Inbound Sea-Teachers Batch Period Outbound Inbound 2 17 Jul- 19 Aug 2016 6 11 3 15 Jan- 13 Feb 2017 6 13 4 7 Aug- 5 Sep 2017 12 16 5 22 Jan- 21 Feb 2018 3 18 6 11 Aug- 14 Sep 2018 17 17 7 11 Jan- 14 Feb 2019 12 16 8 12 Aug- 15 Sep 2019 11 19 Total 67 110 CRRU Universities Partners (Outbound) PGRI University of Universitas Tanjungpura Indonesia University of Edu Tidar University Yogyakarta Benguet State University Islamic University of Indon Philippine Normal Unive Universitas Ahmad Dahlan Cavite State University Islamic University of Kalima Saint Mary’s University Universitas Islam Kalimantan Far Eastern University Lambung Mangkurat Unive Sebelas Maret University Universitas Negeri Makassar Universitas Sarjanawiyata Halu Oleo University Pakuan University Tadulako University Tamansiswa Iloilo Science And Technology University of Mercu Buana Pangasinan State UniversityTarlac Agricultural Unive University Yogyakarta University of San Jose- Recoletos Yogyakarta State University 26Universities(9 Phillipines17Indonesia) CRRU Universities Partners (Inbound) Indonesia University of Education Miriam College State University of Makassar Universitas Muhammadiyah Purwokerto Ahmad Dahlan University Nusa Cendana University State University of Malang Universitas Negeri -

Pengumuman-FER-FAC
MINISTRY OF RESEARCH, TECHNOLOGY AND HIGHER EDUCATION DIRECTORATE GENERAL OF LEARNING AND STUDENT AFFAIRS THE SUPPORT TO THE DEVELOPMENT OF HIGHER EDUCATION IDB Loan No. IND-0168 and SFD Loan No.9/612 Jl. Tirtayasa Raya No.12, Kebayoran Baru, Jakarta Selatan Telp: (021) 29305072/74 Fax: (021) 29305072; E-mail: [email protected] PENGUMUMAN HASIL EVALUASI PROPOSAL FINANSIAL PENGADAAN FINANCIAL AUDITING CONSULTANT (FAC) UNTUK SAMRATULANGI UNIVERSITY, STATE UNIVERSITY OF GORONTALO, TANJUNGPURA UNIVERSITY, LAMBUNG MANGKURAT UNIVERSITY, STATE UNIVERSITY OF SURABAYA, AND STATE UNIVERSITY OF YOGYAKARTA THE SUPPORT TO THE DEVELOPMENT OF HIGHER EDUCATION PROJECT TAHUN ANGGARAN 2017 Nomor: 809/PP.FAC.IDB.PMU/IX/2017 Berdasarkan hasil evaluasi Panitia Seleksi terhadap proposal finansial tanggal 7 April 2017 serta No Objection Letter (NOL) dari Islamic Development Bank (IDB) nomor HDE/EDU/867 tanggal 10 September 2017, dengan ini kami umumkan bahwa perusahaan dengan penawaran terendah pada evaluasi proposal finansial untuk: Nama Pekerjaan : Pengadaan Financial Auditing Consultant (FAC) untuk Universitas Sam Ratulangi, Universitas Negeri Gorontalo, Universitas Tanjungpura, Universitas Lambung Mangkurat, Universitas Negeri Surabaya, dan Universitas Negeri Yogyakarta, The Support to The Development of Higher Education Project Metoda : Least-Cost Selection (LCS) Sumber Dana : DIPA Direktorat Jenderal Pembelajaran dan Kemahasiswaan Kementerian Riset, Teknologi, dan Pendidikan Tinggi Tahun Anggaran 2017 yang bersumber dari pinjaman Islamic Development Bank (IDB Loan No. IND-0168) Adalah sebagai berikut: Nama Perusahaan Alamat Nilai Penawaran KAP Heliantono & TB. Simatupang Kav.10, Jakarta Rp 2,330,000,000,- Rekan Demikian disampaikan, atas partisipasinya diucapkan terima kasih. Jakarta, 11 September 2017 Panitia Seleksi . -

Kalimantan Barat, Indonesia - Sarawak, Malaysia “Head to Head”
Borderlands and Tourism Development in Kalimantan Island: Kalimantan Barat, Indonesia - Sarawak, Malaysia “Head to Head” Syaiful Muazir Department of Architecture, Tanjungpura University, Pontianak, Indonesia [email protected] Horng-Chang Hsieh Department of Urban Planning, National Cheng Kung University, Tainan, Taiwan [email protected] As part of the regions that share lands and boundaries, Sarawak-Malaysia and Kalimantan Barat- Indonesia have respective characteristics in the borderlands treatment. This paper aims to explore and compare the border areas between two nations, ranging from the borderlands profiles, (tourism) development and planning, and their opportunities. Besides, this paper also wants to highlight tourism development as one of the strategies. This paper uses documentary research to seek “text and context” by narratively describing and compiling the documents. From the discussion, it can be seen that both regions have different characteristics of conditions and problems. For tourism development, different approaches are also “colored” with timing, momentum, and priorities; as influenced and supported by the national plans. Tourism in the borderlands is the impetus in which each regions has already been aware of this potential because of the traction of informal flow, accessibility, supply, and demand. Furthermore, we argue that if tourism has exist at the first place, in the future, by its backward linkages, it will encourage more physical and economic development and cooperation between nations. Keywords: borderland, tourism development, Sarawak, Kalimantan Barat 1. INTRODUCTION diplomatic relationship. There is a concept of “blood-brothers” that can be tracked through Indonesia and Malaysia have a long historical ideology, history, commonalities of race, ethnicity, background of bilateral relations, as well as on the and cultural forms. -

Analysis of River Flow Regime Changes Related to Water Availability on the Kapuas River, Indonesia†
IRRIGATION AND DRAINAGE Irrig. and Drain. (2017) Published online in Wiley Online Library (wileyonlinelibrary.com) DOI: 10.1002/ird.2103 ANALYSIS OF RIVER FLOW REGIME CHANGES RELATED TO WATER AVAILABILITY ON THE KAPUAS RIVER, INDONESIA† HENNY HERAWATI1*, S. SURIPIN2, S. SUHARYANTO2 AND TIA HETWISARI3 1Civil Engineering Department, Tanjungpura University, Pontianak, Indonesia 2Civil Engineering Department, Diponegoro University, Semarang, Indonesia 3Directorate General of Human Settlement, Ministry of Public Works, Semarang, Indonesia ABSTRACT Rivers are major sources of fresh water, which is a basic need in society. The availability of water is affected by river basin characteristics such as rainfall and land cover type. Along with population and economy growth, there has been a change in land use that leads to changes in land cover types, thereby changing the river flow regime and affecting the availability of water in the river. Thus it is necessary to analyse water flow regime changes in the river to determine water availability for proper action plans in the future. Studies on these aspects were conducted in the Kapuas River Basin with an area of about 100 000 km2, in West Kalimantan, Indonesia. The study was conducted by analysing the trend of annual rainfall and change of land cover during the last three decades, by analysing land cover types and comparing the availability of water in the Kapuas River from measurement results in 2002 and 2012. The results showed that the rainfall trend had increased 4.3% over the last 30 years. The changes in the hydrological aspects of the study area have altered the river flow regime of Kapuas River within the last 30 years and have influenced river water availability. -

Strengthening the Centre of Research Excellence at Universitas Ma Chung at National and International Level
Strengthening the Centre of Research Excellence at Universitas Ma Chung at national and international Level Leenawaty Limantara Abstract Higher education reforms happening in Europe, Africa, South-East Asia and all over the world demand changes at higher education institutions. The reforms are a consequence of globalization, expanding access to higher education, new technol- ogies (computer, internet), new concept in public management, formidable com- petition, and convergence of higher education policies. Universitas Ma Chung as a new university in the midst of competition of 3,673 state and private universities in Indonesia, and universities around the globe, implements higher education man- agement strategies employing a variety of approaches. This paper addresses one of its management strategies using the Project Action Plan (PAP) to strengthen the University Centre of Excellence aiming at sharpening and brightening the colour and excellence in the university research sector at national and international level through four activities which are expected to make impact internally, nationally, and globally. They are (a) a new grand design of the CORE Ma Chung Research Centre for Photosynthetic Pigments (MRCCP) in biotechnology, (b) creating the foundation of excellent research atmosphere, (c) strengthening the existing col- laboration and the position in the international consortium, and (d) creating the national consortium in the core area of research. The result of the PAP implemen- tation over eight months shows that (1) a project action plan is a reliable tool for initiating, planning, implementing, controlling, and evaluating a project to help an organization in making a project succeed by determining the time, place, person in charge (PIC), personnel involved, support needed, and is a measuring instrument to gauge detailed success with clear milestones. -

List of English and Native Language Names
LIST OF ENGLISH AND NATIVE LANGUAGE NAMES ALBANIA ALGERIA (continued) Name in English Native language name Name in English Native language name University of Arts Universiteti i Arteve Abdelhamid Mehri University Université Abdelhamid Mehri University of New York at Universiteti i New York-ut në of Constantine 2 Constantine 2 Tirana Tiranë Abdellah Arbaoui National Ecole nationale supérieure Aldent University Universiteti Aldent School of Hydraulic d’Hydraulique Abdellah Arbaoui Aleksandër Moisiu University Universiteti Aleksandër Moisiu i Engineering of Durres Durrësit Abderahmane Mira University Université Abderrahmane Mira de Aleksandër Xhuvani University Universiteti i Elbasanit of Béjaïa Béjaïa of Elbasan Aleksandër Xhuvani Abou Elkacem Sa^adallah Université Abou Elkacem ^ ’ Agricultural University of Universiteti Bujqësor i Tiranës University of Algiers 2 Saadallah d Alger 2 Tirana Advanced School of Commerce Ecole supérieure de Commerce Epoka University Universiteti Epoka Ahmed Ben Bella University of Université Ahmed Ben Bella ’ European University in Tirana Universiteti Europian i Tiranës Oran 1 d Oran 1 “Luigj Gurakuqi” University of Universiteti i Shkodrës ‘Luigj Ahmed Ben Yahia El Centre Universitaire Ahmed Ben Shkodra Gurakuqi’ Wancharissi University Centre Yahia El Wancharissi de of Tissemsilt Tissemsilt Tirana University of Sport Universiteti i Sporteve të Tiranës Ahmed Draya University of Université Ahmed Draïa d’Adrar University of Tirana Universiteti i Tiranës Adrar University of Vlora ‘Ismail Universiteti i Vlorës ‘Ismail -

Download Article (PDF)
Advances in Social Science, Education and Humanities Research, volume 422 International Conference on Progressive Education (ICOPE 2019) Profile of Undergraduate Students as Prospective Science Teachers in terms of Science Literacy Tulus Junanto Budiyono Universitas Sebelas Maret Universitas Sebelas Maret Doctoral Program of Educational Faculty of Educational Science, Knowledge Teacher Training and Education Teacher Training and Education Faculty of Sebelas Maret University Faculty of Sebelas Maret University Surakarta, Indonesia Surakarta, Indonesia e-mail: budiyono.staff.uns.ac.id e-mail: [email protected] Nunuk Suryani Muhammad Akhyar Universitas Sebelas Maret Universitas Sebelas Maret Faculty of Educational Science, Faculty of Educational Science, Teacher Training and Education Teacher Training and Education Faculty of Sebelas Maret University Faculty of Sebelas Maret University Surakarta, Indonesia Surakarta, Indonesia e-mail: [email protected] e-mail: [email protected] Abstract— This study aims to describe the profile of the able to meet the needs of life in various situations, namely the scientific literacy skills of undergraduate students at ability of scientific literacy. Tanjungpura University, Pontianak. The research method used is a descriptive method to describe the initial ability of Nowadays, science literacy is a discussion in the world of scientific literacy of undergraduate students as prospective education. Many developed and developing countries have science teachers, especially in chemistry. The research subjects made science literacy the goal of science learning. Science were undergraduate students in the chemistry education study literacy is the ability to understand science, communicate program. The research sample uses a purposive sampling science, and apply the ability of science to solve problems. -
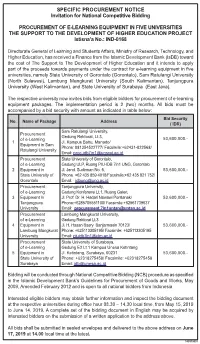
Pengumuman E-Learning.Pdf
SPECIFIC PROCUREMENT NOTICE Invitation for National Competitive Bidding PROCUREMENT OF E-LEARNING EQUIPMENT IN FIVE UNIVERSITIES THE SUPPORT TO THE DEVELOPMENT OF HIGHER EDUCATION PROJECT Istisna’a No.: IND-0168 Directorate General of Learning and Students Affairs, Ministry of Research, Technology, and Higher Education, has received a Finance from the Islamic Development Bank (IsDB) toward the cost of The Support to The Development of Higher Education and it intends to apply part of the proceeds towards payments under the contract for e-learning equipment in five universities, namely State University of Gorontalo (Gorontalo), Sam Ratulangi University (North Sulawesi), Lambung Mangkurat University (South Kalimantan), Tanjungpura University (West Kalimantan), and State University of Surabaya (East Java). The respective university now invites bids from eligible bidders for procurement of e-learning equipment packages. The implementation period is 2 (two) months. All bids must be accompanied by a bid security with amount as indicated in table below: Bid Security No. Name of Package Address ( IDR) Sam Ratulangi University, Procurement Gedung Rektorat, Lt.3, of e-Learning 53,600,000.- 1 Jl. Kampus Bahu, Manado/ Equipment in Sam Phone: 081354522777/ Facsimile:+62431-822568/ Ratulangi University Email: [email protected] Procurement State University of Gorontalo, of e-Learning Gedung ULP, Ruang PIU-IDB 7in1 UNG, Gorontalo 2 Equipment in Jl. Jend. Sudirman No. 6, 53,600,000.- State University of Phone +62 435 859 4018/Facsimile:+62 435 821 752/ Gorontalo Email: [email protected] Procurement Tanjungpura University, of e-Learning Gedung Konferensi Lt.1, Ruang Galeri, 3 Equipment in Jl. Prof. Dr.