Revision of the Giant Geckos of New Caledonia (Reptilia: Diplodactylidae: Rhacodactylus)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
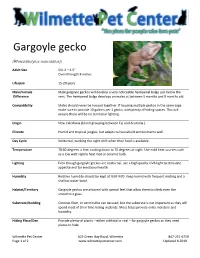
Gargoyle Gecko
Gargoyle gecko (Rhacodactylus auriculatus)) Adult Size SVL 4 – 4.5” Overall length 8 inches Lifespan 15-20 years Male/Female Male gargoyle geckos will develop a very noticeable hemipenal bulge just below the Difference vent. The hemipenal bulge develops on males at between 5 months and 9 months old. Compatibility Males should never be housed together. If housing multiple geckos in the same cage make sure to provide 10 gallons per 1 gecko, with plenty of hiding spaces. This will ensure there will be no territorial fighting. Origin New Caledonia (Island grouping between Fiji and Australia.) Climate Humid and tropical jungles, but adapts to household environments well. Day Cycle Nocturnal, working the night shift when their food is available. Temperature 78-82 degrees is fine, cooling down to 70 degrees at night. Use mild heat sources such as a low watt reptile heat mat or ceramic bulb. Lighting Even though gargoyle geckos are nocturnal, use a high quality UVA light to stimulate appetite and for emotional health. Humidity Relative humidity should be kept at %50-%70. Keep humid with frequent misting and a shallow water bowl. Habitat/Territory Gargoyle geckos are arboreal with special feet that allow them to climb even the smoothest glass. Substrate/Bedding Coconut fiber, or vermiculite can be used, but the substrate is not important as they will spend most of their time hiding in plants. Moss helps provide extra moisture and humidity. Hiding Place/Den Provide plenty of plants – either artificial or real – for gargoyle geckos as they need places to hide. Wilmette Pet Center 625 Green Bay Road, Wilmette 847-251-6750 Page 1 of 2 www.wilmettepetcenter.com Updated 4.2019 Cage Type Ten gallon aquariums or critter cages with screen tops work well for gargoyle geckos. -

Crested Gecko by Catherine Love, DVM Updated 2021
Crested Gecko By Catherine Love, DVM Updated 2021 Natural History Rhacodactylus ciliatus, more recently re-classified as Correlophus ciliatus, is a species of arboreal lizard native to New Caledonia. Until 1994, crested geckos were thought to be extinct in the wild. Their population was re-discovered, and although export is no longer allowed, this species has thrived in captivity and is readily available in the US pet trade. Cresties are crepuscular (active at dawn and dusk), and spend most of their day in shrubs or in trees. They get their name from the eyelash-like appendages that form the crests above their eyes. Unlike most geckos that are carnivorous, these animals are omnivorous, and primarily frugivores (fruit eaters). One of the biggest threats to wild crested geckos is the introduced little fire ant that not only swarms the geckos in large numbers, but also competes for arthropod prey. Crested geckos are considered “vulnerable” by the IUCN. Characteristics and Behavior As with most geckos, cresties do not have eyelids, so they keep their eyes moist by licking them. Cresties are also able to climb vertical surfaces using tiny hairs on their feet called setae. Their tails are partially prehensile, though not to the same extent as a chameleon, and they possess tail autotomy (they can drop their tails). Unlike leopard geckos, when a crestie drops their tail, it doesn’t grow back. However, they don’t store fat in their tails so it is generally not as detrimental for a crestie to drop their tail, and the majority of wild cresties end up doing so. -

THE UNDERCROFT We Must Expire in Hope{ of Resurrection to Life Again
THE UNDERCROFT We must Expire in hope{ of Resurrection to Life Again 1 Editorial Illu{tration{ Daniel Sell Matthew Adams - 43 Jeremy Duncan - 51, 63 Anxious P. - 16 2 Skinned Moon Daughter Cedric Plante - Cover, 3, 6, 8, 9 Benjamin Baugh Sean Poppe - 28 10 101 Uses of a Hanged Man Barry Blatt Layout Editing & De{ign Daniel Sell 15 The Doctor Patrick Stuart 23 Everyone is an Adventurer E{teemed Con{ultant{ Daniel Sell James Maliszewski 25 The Sickness Luke Gearing 29 Dead Inside Edward Lockhart 41 Cockdicktastrophe Chris Lawson 47 Nine Summits and the Matter of Birth Ezra Claverie M de in elson Ma ia S t y a it mp ic of Authent Editorial Authors live! Hitler writes a children’s book, scratches his name off and hides it among the others. Now children grow up blue and blond, unable to control their minds, the hidden taint in friendly balloons and curious caterpillars wormed itself inside and reprogrammed them into literary sleeper agents ready to steal our freedom. The swine! We never saw it coming. If only authors were dead. But if authors were dead you wouldn’t be able to find them, to sit at their side and learn all they had to say on art and beautiful things, on who is wrong and who is right, on what is allowed and what is ugly and what should be done about this and that. We would have only words and pictures on a page, if we can’t fill in the periphery with context then how will we understand anything? If they don’t tell us what to enjoy, how to correctly enjoy it, then how will we know? Who is right and who is mad? Whose ideas are poisonous and wrong? Call to arms! Stop that! Help me! We must slather ourselves in their selfish juices. -

Keystone Species: the Concept and Its Relevance for Conservation Management in New Zealand
Keystone species: the concept and its relevance for conservation management in New Zealand SCIENCE FOR CONSERVATION 203 Ian J. Payton, Michael Fenner, William G. Lee Published by Department of Conservation P.O. Box 10-420 Wellington, New Zealand Science for Conservation is a scientific monograph series presenting research funded by New Zealand Department of Conservation (DOC). Manuscripts are internally and externally peer-reviewed; resulting publications are considered part of the formal international scientific literature. Titles are listed in the DOC Science Publishing catalogue on the departmental website http:// www.doc.govt.nz and printed copies can be purchased from [email protected] © Copyright July 2002, New Zealand Department of Conservation ISSN 11732946 ISBN 047822284X This report was prepared for publication by DOC Science Publishing, Science & Research Unit; editing by Lynette Clelland and layout by Ruth Munro. Publication was approved by the Manager, Science & Research Unit, Science Technology and Information Services, Department of Conservation, Wellington. CONTENTS Abstract 5 1. Introduction 6 2. Keystone concepts 6 3. Types of keystone species 8 3.1 Organisms controlling potential dominants 8 3.2 Resource providers 10 3.3 Mutualists 11 3.4 Ecosystem engineers 12 4. The New Zealand context 14 4.1 Organisms controlling potential dominants 14 4.2 Resource providers 16 4.3 Mutualists 18 4.4 Ecosystem engineers 19 5. Identifying keystone species 20 6. Implications for conservation management 21 7. Acknowledgements 22 8. References 23 4 Payton et al.Keystone species: the concept and its relevance in New Zealand Keystone species: the concept and its relevance for conservation management in New Zealand Ian J. -

Integrative and Comparative Biology Integrative and Comparative Biology, Pp
Integrative and Comparative Biology Integrative and Comparative Biology, pp. 1–17 doi:10.1093/icb/icz006 Society for Integrative and Comparative Biology SYMPOSIUM 2019 April 28 on user Cities Twin - Minnesota of by University https://academic.oup.com/icb/advance-article-abstract/doi/10.1093/icb/icz006/5381544 from Downloaded Evolution of the Gekkotan Adhesive System: Does Digit Anatomy Point to One or More Origins? Anthony P. Russell1,* and Tony Gamble†,‡,§ *Department of Biological Sciences, University of Calgary, 2500 University Drive NW, Calgary, Alberta, Canada T2N 1N4; †Department of Biological Sciences, Marquette University, Milwaukee, WI 53201, USA; ‡Bell Museum of Natural History, University of Minnesota, Saint Paul, MN 55113, USA; §Milwaukee Public Museum, Milwaukee, WI 53233, USA From the symposium “The path less traveled: Reciprocal illumination of gecko adhesion by unifying material science, biomechanics, ecology, and evolution” presented at the annual meeting of the Society of Integrative and Comparative Biology, January 3–7, 2019 at Tampa, Florida. 1E-mail: [email protected] Synopsis Recently-developed, molecularly-based phylogenies of geckos have provided the basis for reassessing the number of times adhesive toe-pads have arisen within the Gekkota. At present both a single origin and multiple origin hypotheses prevail, each of which has consequences that relate to explanations about digit form and evolutionary transitions underlying the enormous variation in adhesive toe pad structure among extant, limbed geckos (pygopods lack pertinent features). These competing hypotheses result from mapping the distribution of toe pads onto a phylo- genetic framework employing the simple binary expedient of whether such toe pads are present or absent. -

Two New Species of the Genus Bavayia (Reptilia: Squamata: Diplodactylidae) from New Caledonia, Southwest Pacific!
Pacific Science (1998), vol. 52, no. 4: 342-355 © 1998 by University of Hawai'i Press. All rights reserved Two New Species of the Genus Bavayia (Reptilia: Squamata: Diplodactylidae) from New Caledonia, Southwest Pacific! AARON M. BAUER, 2 ANTHONY H. WHITAKER,3 AND Ross A. SADLIER4 ABSTRACT: Two new species ofthe diplodactylid gecko Bavayia are described from restricted areas within the main island ofNew Caledonia. Both species are characterized by small size, a single row of preanal pores, and distinctive dorsal color patterns. One species is known only from the endangered sclerophyll forest of the drier west coast of New Caledonia, where it was collected in the largest remaining patch of such habitat on the Pindai" Peninsula. The second species occupies the maquis and adjacent midelevation humid forest habitats in the vicinity of Me Adeo in south-central New Caledonia. Although relation ships within the genus Bavayia remain unknown, the two new species appear to be closely related to one another. BAVAYIA IS ONE OF THREE genera of carpho tion of the main island. The two most wide dactyline geckos that are endemic to the New spread species, B. cyclura and B. sauvagii, are Caledonian region. Seven species are cur both probably composites of several mor rently recognized in the genus (Bauer 1990). phologically similar, cryptic sibling species. Three of these, B. crassicollis Roux, B. cy Recent field investigations on the New Cale clura (Giinther), and B. sauvagii (Boulenger), donian mainland have revealed the presence are relatively widely distributed, with popula of two additional species of Bavayia. Both tions on the Isle of Pines (Bauer and Sadlier are small, distinctively patterned, and appar 1994) and the Loyalty Islands (Sadlier and ently restricted in distribution. -

RVC Exotics Service CRESTED GECKO CARE
RVC Exotics Service Royal Veterinary College Royal College Street London NW1 0TU T: 0207 554 3528 F: 0207 388 8124 www.rvc.ac.uk/BSAH CRESTED GECKO CARE The Crested gecko (Rhacodactylus ciliatus) originates from the islands of New Caledonia where the species was believed to be extinct until 1994 when it was subsequently rediscovered. In its natural environment, it can be found resting in rainforests, sleeping in trunk hollows or leaf litter, and only becomes active at night. Similar to other geckos, it can shed its tail as a defence mechanism (autotomy), but unlike others it lacks an ability to regenerate the tail, so care should be taken while handling. Geckos may live 10-15 years if looked after correctly. HOUSING • As large a vivarium as possible should be provided to enable room for exercise, and a thermal gradient to be created along the length of the tank (hot to cold). Wooden or fibreglass vivaria are ideal as this provides the lizard with some visual security and ventilation can be provided at lizard level. • Good ventilation is required and additional ventilation holes may need to be created. • Hides are required to provide some security. Artificial plants, cardboard boxes, plant pots, logs or commercially available hides can be used. They should be placed both at the warm and cooler ends of the tank. One hide should contain damp moss, kitchen towel or vermiculite to provide a humid environment for shedding. • Substrates suitable for housing lizards include newspaper, Astroturf and some of the commercially available substrates. It is important that the substrates either cannot be eaten, or if they are, do not cause blockages as this can prove fatal. -

A New Locality for Correlophus Ciliatus and Rhacodactylus Leachianus (Sauria: Diplodactylidae) from Néhoué River, Northern New Caledonia
Herpetology Notes, volume 8: 553-555 (2015) (published online on 06 December 2015) A new locality for Correlophus ciliatus and Rhacodactylus leachianus (Sauria: Diplodactylidae) from Néhoué River, northern New Caledonia Mickaël Sanchez1, Jean-Jérôme Cassan2 and Thomas Duval3,* Giant geckos from New Caledonia (Pacific Ocean) We observed seven native gecko species: Bavayia are charismatic nocturnal lizards. This paraphyletic (aff.) cyclura (n=1), Bavayia (aff.) exsuccida (n=1), group is represented by three genera, Rhacodactylus, Correlophus ciliatus (n=1), Dierrogekko nehoueensis Correlophus and Mniarogekko, all endemic to Bauer, Jackman, Sadlier and Whitaker, 2006 (n=1), New Caledonia (Bauer et al., 2012). Rhacodactylus Eurydactylodes agricolae Henkel and Böhme, 2001 leachianus (Cuvier, 1829) is largely distributed on the (n=1), Mniarogekko jalu Bauer, Whitaker, Sadlier and Grande Terre including the Île des Pins and its satellite Jackman, 2012 (n=1) and Rhacodactylus leachianus islands, whereas Correlophus ciliatus Guichenot, 1866 (n=1). Also, the alien Hemidactylus frenatus Dumeril is mostly known in the southern part of the Grande and Bibron, 1836 (n=3) has been sighted. The occurrence Terre, the Île des Pins and its satellite islands (Bauer of C. ciliatus and R. leachianus (Fig. 2 and 3) represent et al., 2012). Here, we report a new locality for both new records for this site. Both gecko species were species in the north-western part of Grande Terre, along observed close to the ground, at a height of less than the Néhoué River (Fig. 1). 1.5 m. The Néhoué River is characterized by gallery forests It is the first time that R. leachianus is recorded in the growing on deep alluvial soils. -

Thermal Ecology and Habitat Utilization of Rhacodactylus Leachianus from New Caledonia (Squamata: Diplodactylidae)
SALAMANDRA 54(2)Thermal117–122 ecology and15 Mayhabitat 2018 utilizationISSN of 0036–3375 Rhacodactylus leachianus from New Caledonia Thermal ecology and habitat utilization of Rhacodactylus leachianus from New Caledonia (Squamata: Diplodactylidae) Peter Sound1, Friedrich Wilhelm Henkel2, Christian Langner3 & Alfred Seitz1 1) Zoologisches Institut, Abteilung V Populationsbiologie, Universität Mainz, Saarstr. 21, 55099 Mainz, Germany 2) Frielinger Weg 25a, 59174 Kamen, Germany 3) Alstätte 23, 48727 Billerbeck, Germany Corresponding author: Peter Sound, e-mail: [email protected] Received: 24 March 2015 Accepted: 6 February 2018 by Stefan Lötters Abstract. Five specimens (2 ♀, 3 ♂) of the gecko species Rhacodactylus leachianus were radio-tracked for 14 days on Île de Bayonnaise, New Caledonia. Using thermo-sensitive transmitters, specimens were mapped continuously every hour. All movements, behavioural expressions and core temperatures were manually recorded, while data loggers simultaneously recorded the temperatures of microhabitat structures. In an additional trial, all geckos captured on the island were marked with Passive Integrated Transponders (PITs): Twenty-two adults (13 ♀, 9 ♂) and one subadult were marked in total. The mapping of R. leachianus on this island was repeated during 2005, 2007 and 2013. Average core temperatures of 23.4°C (♀) and 24.1°C (♂) were recorded ranging of 18.2–32.0°C (♀) and 18.0–32.2°C (♂), respectively. No sex-dependent differences were noted, but a clear relation to the local air temperature was found. Gravid females showed higher core temperatures (by up to 3°C) during oviposition. The test for equality of variances showed differences between the core temperatures of both sexes and the air temperature, indicating the ability of both sexes for active thermoregulation. -

Reptile Vacation – Visiting Our Homelands by Chelsea Holden
Reptile Vacation – Visiting Our Homelands By Chelsea Holden It was the middle of winter, and the reptiles in the lab were getting restless. They were all tired of the snow and cold and wanted to go on a vacation. But where would they go? All of the reptiles thought that their homeland would be the best place to visit. The bearded dragons, Frieda and Freddie, were the first to speak up. “Let’s go to Australia!” said Freddie. “We can go and be warm with trees, rocks, and maybe some sand. The perfect spot would have to be Queensland. While basking in the sun, we’ll have it made!” “And if it gets too hot, we’ll cool off in the shade!” added Frieda. The Rankin’s dragons also agreed. “To Queensland! To Queensland!” they shouted with glee. The frilled dragons had other ideas. “Australia would be nice,” said Villain, “but we think it’d be better if we went somewhere warm and wetter. And we want to go hide in a tree!” The other frilled dragons could only agree. From across the room, there came a rustling sound. It was the sand boa, poking its head out of the ground! “Sand sounds lovely,” it hissed. “It’s the desert for me. African sands are the place to be! I’d burrow my way into the warm sands and take in the sights of the Great Pyramids.” The green tree python piped in: “You can keep your sand! It’s the trees for me! The rainforests of New Guinea are the place to be! I’ll lounge on the tree branches and hide in the canopies.” Overhearing the excitement, the Leachies joined in: “We know where some of the geckos would love to go. -

Species Boundaries, Biogeography, and Intra-Archipelago Genetic Variation Within the Emoia Samoensis Species Group in the Vanuatu Archipelago and Oceania" (2008)
Louisiana State University LSU Digital Commons LSU Doctoral Dissertations Graduate School 2008 Species boundaries, biogeography, and intra- archipelago genetic variation within the Emoia samoensis species group in the Vanuatu Archipelago and Oceania Alison Madeline Hamilton Louisiana State University and Agricultural and Mechanical College, [email protected] Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_dissertations Recommended Citation Hamilton, Alison Madeline, "Species boundaries, biogeography, and intra-archipelago genetic variation within the Emoia samoensis species group in the Vanuatu Archipelago and Oceania" (2008). LSU Doctoral Dissertations. 3940. https://digitalcommons.lsu.edu/gradschool_dissertations/3940 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Doctoral Dissertations by an authorized graduate school editor of LSU Digital Commons. For more information, please [email protected]. SPECIES BOUNDARIES, BIOGEOGRAPHY, AND INTRA-ARCHIPELAGO GENETIC VARIATION WITHIN THE EMOIA SAMOENSIS SPECIES GROUP IN THE VANUATU ARCHIPELAGO AND OCEANIA A Dissertation Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Doctor of Philosophy in The Department of Biological Sciences by Alison M. Hamilton B.A., Simon’s Rock College of Bard, 1993 M.S., University of Florida, 2000 December 2008 ACKNOWLEDGMENTS I thank my graduate advisor, Dr. Christopher C. Austin, for sharing his enthusiasm for reptile diversity in Oceania with me, and for encouraging me to pursue research in Vanuatu. His knowledge of the logistics of conducting research in the Pacific has been invaluable to me during this process. -

Gargoyle Gecko Reptile - a Cold-Blooded Vertebrate with Scaly Skin
Glossary Gargoyle Gecko Reptile - A cold-blooded vertebrate with scaly skin. The gargoyle gecko is a nocturnal, arboreal lizard Amphibian - A cold-blooded vertebrate that begins life as that makes for a great companion. They come in an aquatic animal and grows into a terrestrial adult with many colourations (morphs) and are best recognised lungs. for their big eyes. They are closely related to the Terrestrial - A ground dwelling animal. crested gecko. Males cannot be kept together as Arboreal - An animal that lives in trees. they are aggressive to one another but females can Diurnal - Awake in the day. Gargoyle be housed together. If keeping males and females Nocturnal- Awake during the night. together, it is best to have minimal 2 females to 1 UVB - Ultraviolet radiaton. Gecko male. They can live between 15 to 20 years. Colubrid - A family of snakes. Hybrid - Offspring from animals of different species. Morph - Colourations created due to genetics. Musk - Unpleasant odour released when an animal is stressed or feels threatened. Live plants are only available on special order If you require any further information, please ask our pet care advisors who will be very happy to help. Opening Times Monday - Saturday: 9am - 6pm Sunday: 9.30am - 4pm Chessington Garden Centre Leatherhead Road, Chessington, Surrey, KT9 2NG Care & Advice Sheet Tel: 01372 725 638 Email: [email protected] Web: www.chessingtongardencentre.co.uk Please recycle me once you’ve nished reading. Inspiration for your Home & Garden Substrate & Furnishings Food & Water Substrates that maintain high humidity are Gargoyle geckos are fruit eating geckos. The recommended such as humus bricks or coco fibre.