Death by Smallpox in 18Th and 19Th C. South Africa
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Anglo-Boer War, a Welsh Hospital in South Africa
24/05/2015 9:00 AM http://samilitaryhistory.org/vol123sw.html The South African Military History Society Die Suid-Afrikaanse Krygshistoriese Vereniging Military History Journal Vol 12 No 3 - June 2002 THE ANGLO-BOER WAR A WELSH HOSPITAL IN SOUTH AFRICA SA Watt, Pietermaritzburg The Welsh Hospital was one of a number of private initiatives in the medical services that were accepted and used by the British Government during the Anglo Boer War (1899-1902). It was organised by Professor Alfred W Hughes, assisted by a committee elected from the men and women of Wales. Funds amounting to £12 000 were acquired by subscription from the citizens of Wales and Welshmen residing outside the country. According to a Report by the Central British Red Cross Committee on the Voluntary Organisations in the Aid of the Sick and Wounded during the South African War, the personnel originally comprised three senior surgeons, two assistant surgeons, eight medical students and dressers, ten nursing sisters, two maids, 48 orderlies, cooks, and stretcher bearers. The medal roll lists 44 staff (W A Morgan, 1975, p 12). With them was the matron, Marion Lloyd. One of the senior surgeons was Professor Thomas Jones, who was a professor of surgery at Owen's College, Manchester, England (Report by the CBRCC, 1902; British Medical Journal, p 250). The personnel and equipment under the command of Major T W Cockerill embarked from Southampton on the Canada, 14 April 1900. The passage and freight was provided by the government. The stores, subsequently sent out, were shipped at the expense of the organisers. -

Botshabelo and Thaba Nchu CRDP
Kopanong Ward # 41602001 Comprehensive Rural Development Program Status Quo Report CHIEF DIRECTORATE: SPATIAL PLANNING AND INFORMATION July 25, 2011 Authored by: SPI Free State TABLE OF CONTENTS 1. INTRODUCTION ........................................................................................................ 3 1.2. OBJECTIVE OF THE STUDY ........................................................................................... 3 1.3. BACKGROUND ........................................................................................................ 3 2. RESEARCH DESIGN ..................................................................................................... 4 2.1. PROBLEM STATEMENT ....................................................................................... 4 2.2. METHODOLOGY .............................................................................................. 5 2.2.1. DEFINITION OF RURAL AREAS (OECD) ............................................................... 7 3. STUDY AREA .............................................................................................................. 8 3.1. PROVINCIAL CONTEXT ...................................................................................... 8 3.2. DISTRICT CONTEXT .......................................................................................... 9 3.3. LOCAL CONTEXT ........................................................................................... 10 3.4. PILOT SITE .................................................................................................. -

A Spirituality of Mercy in a Time of War
Religion &Theology Religion & Theology 18 (2011) 147–172 brill.nl/rt A Spirituality of Mercy in a Time of War Pieter G. R. de Villiers Department of Old and New Testament, University of the Free State, P.O. Box 339, Bloemfontein 9300, Republic of South Africa [email protected] Abstract This article discusses the diary of the Rev. A. D. Luckhoff, written during the South African War in the infamous Bethulie concentration camp from the perspective of his Spirituality of mercy. It analyses the diary in terms of mercy as giving love, as charitableness and as respect for human dignity. It then investigates mercy in the diary as compassion and vulnerability, before it con- cludes by discussing how Lukchoff ’s faith in providence, his awareness of a divine call, his inner space of homeliness and love empower mercy. Keywords mercy, vulnerability, compassion, spirituality, A. D. Luckhoff 1. Introduction One of the best-known personalities in the Dutch Reformed church of the early twentieth century was the Rev. Abraham D. Luckhoff. Born in 1874 from a colourful family of pastors from Rhenish, Wesleyan and Dutch Reformed contexts,1 he is first and foremost remembered for his lifelong work in the Dutch Reformed Church’s welfare department for a period of 27 years (1916–1943). His upliftment of communities made a lasting impression on the church, his community and even the country to the extent that his advice on matters of upliftment was often asked by state departments, whilst he was 1 Cf. F. Pretorius’ introduction to A. D. Luckhoff, Woman’s Endurance (Pretoria: Protea, 2006 [1904]), iii–ix. -

Thesis Hum 2007 Mcdonald J.Pdf
The copyright of this thesis vests in the author. No quotation from it or information derived from it is to be published without full acknowledgement of the source. The thesis is to be used for private study or non- commercial research purposes only. Published by the University of Cape Town (UCT) in terms of the non-exclusive license granted to UCT by the author. University of Cape Town 'When Shall These Dry Bones Live?' Interactions between the London Missionary Society and the San Along the Cape's North-Eastern Frontier, 1790-1833. Jared McDonald MCDJAR001 A dissertation submitted in fulfillment of the requirements for the award of the degree of Master of Arts in Historical Studies Faculty of the Humanities University of Cape Town 2007 University of Cape Town COMPULSORY DECLARATION This work has not been previously submitted in whole, or in part, for the award of any degree. It is my own work. Each significant contribution to, and quotation in, this dissertation from the work, or works, of other people has been attributed, and has been cited and referenced. Signature:_fu""""',_..,"<_l_'"'"-_(......\b __ ~_L ___ _ Date: 7 September 2007 "And the Lord said to me, Prophe,\y to these bones, and scry to them, o dry bones. hear the Word ofthe Lord!" Ezekiel 37:4 Town "The Bushmen have remained in greater numbers at this station ... They attend regularly to hear the Word of God but as yet none have experiencedCape the saving effects ofthe Gospe/. When shall these dry bones oflive? Lord thou knowest. -

Phytosociology of the Upper Orange River Valley, South Africa
PHYTOSOCIOLOGY OF THE UPPER ORANGE RIVER VALLEY, SOUTH AFRICA A SYNTAXONOMICAL AND SYNECOLOGICAL STUDY M.J.A.WERGER PROMOTOR: Prof. Dr. V. WESTHOFF PHYTOSOCIOLOGY OF THE UPPER ORANGE RIVER VALLEY, SOUTH AFRICA A SYNTAXONOMICAL AND SYNECOLOGICAL STUDY PROEFSCHRIFT TER VERKRUGING VAN DE GRAAD VAN DOCTOR IN DE WISKUNDE EN NATUURWETENSCHAPPEN AAN DE KATHOLIEKE UNIVERSITEIT TE NIJMEGEN, OP GEZAG VAN DE RECTOR MAGNIFICUS PROF. MR. F J.F.M. DUYNSTEE VOLGENS BESLUIT VAN HET COLLEGE VAN DECANEN IN HET OPENBAAR TE VERDEDIGEN OP 10 MEI 1973 DES NAMIDDAGS TE 4.00 UUR. DOOR MARINUS JOHANNES ANTONIUS WERGER GEBOREN TE ENSCHEDE 1973 V&R PRETORIA aan mijn ouders Frontiepieae: Panorama drawn by R.J. GORDON when he discovered the Orange River at "De Fraaye Schoot" near the present Bethulie, probably on the 23rd December 1777. I. INTRODUCTION When the government of the Republic of South Africa in the early sixties decided to initiate a comprehensive water development scheme of its largest single water resource, the Orange River, this gave rise to a wide range of basic and applied scientific sur veys of that area. The reasons for these surveys were threefold: (1) The huge capital investment on such a water scheme can only be justified economically on a long term basis. Basic to this is that the waterworks be protected, over a long period of time, against inefficiency caused by for example silting. Therefore, management reports of the catchment area should.be produced. (2) In order to enable effective long term planning of the management and use of the natural resources in the area it is necessary to know the state of the local ecosystems before a major change is instituted. -

Xhariep Magisterial District
!. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. !. XXhhaarriieepp MMaaggiisstteerriiaall DDiissttrriicctt !. !. !. !. !. !. !. !. !. !. TheunissenS ubD istrict !. BARKLY WEST R59 R707 !. ST DEALESVILLE R708 SAPS WINBURG ST R70 !. R370 Lejwelepuitsa SAPS Dealesville R73 Winburg ST ST ST R31 Lejwelepuitsa ST Marquard !. ST LKN12 Boshof !. BRANDFORT Brandfort SAPS !. Soutpan R703 SAPS !. R64 STR64 Magiisteriiall R703 !. ST Sub District Marquard Senekal CAMPBELL ST Kimberley Dealesville ST Ficksburg !. !. !. !. R64 Sub BOSHOF SOUTPAN SAPS KIMBERLEY ST Sub !. !. !. SAPS Diistriict R64 SAPS Brandfort !. SAPS ST R709 Sub District District Sub !. !. Sub ST District Verkeerdevlei MARQUARD Sub District N1 !. SAPS Clocolan !. !. District STR700 KL District -
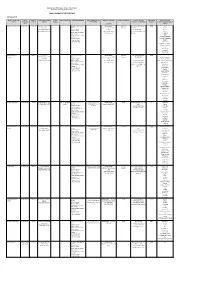
Public Libraries in the Free State
Department of Sport, Arts, Culture & Recreation Directorate Library and Archive Services PUBLIC LIBRARIES IN THE FREE STATE MOTHEO DISTRICT NAME OF FRONTLINE TYPE OF LEVEL OF TOWN/STREET/STREET STAND GPS COORDINATES SERVICES RENDERED SPECIAL SERVICES AND SERVICE STANDARDS POPULATION SERVED CONTACT DETAILS REGISTERED PERIODICALS AND OFFICE FRONTLINE SERVICE NUMBER NUMBER PROGRAMMES CENTER/OFFICE MANAGER MEMBERS NEWSPAPERS AVAILABLE IN OFFICE LIBRARY: (CHARTER) Bainsvlei Public Library Public Library Library Boerneef Street, P O Information and Reference Library hours: 446 142 Ms K Niewoudt Tel: (051) 5525 Car SA Box 37352, Services Ma-Tue, Thu-Fri: 10:00- (Metro) 446-3180 Fair Lady LANGENHOVENPARK, Outreach Services 17:00 Fax: (051) 446-1997 Finesse BLOEMFONTEIN, 9330 Electronic Books Wed: 10:00-18:00 karien.nieuwoudt@mangau Hoezit Government Info Services Sat: 8:30-12:00 ng.co.za Huisgenoot Study Facilities Prescribed books of tertiary Idees Institutions Landbouweekblad Computer Services: National Geographic Internet Access Rapport Word Processing Rooi Rose SA Garden and Home SA Sports Illustrated Sarie The New Age Volksblad Your Family Bloemfontein City Public Library Library c/o 64 Charles Information and Reference Library hours: 443 142 Ms Mpumie Mnyanda 6489 Library Street/West Burger St, P Services Ma-Tue, Thu-Fri: 10:00- (Metro) 051 405 8583 Africa Geographic O Box 1029, Outreach Services 17:00 Architect and Builder BLOEMFONTEIN, 9300 Electronic Books Wed: 10:00-18:00 Tel: (051) 405-8583 Better Homes and Garden n Government Info -

Federal Register/Vol. 74, No. 142/Monday, July 27, 2009/Notices
37000 Federal Register / Vol. 74, No. 142 / Monday, July 27, 2009 / Notices Africa from which citrus fruit is Inspection Service has determined the contact Dr. Patricia L. Foley, Center for authorized for importation into the regulatory review period for NAHVAX® Veterinary Biologics, Policy Evaluation United States, our review of the Marek’s Disease Vaccine and is and Licensing, VS, APHIS, 510 South information presented by the Republic publishing this notice of that 17th Street, Suite 104, Ames, IA 50010; of South Africa in support of its request determination as required by law. We phone (515) 232–5785, fax (515) 232– is examined in a commodity import have made this determination in 7120. evaluation document (CIED) titled response to the submission of an SUPPLEMENTARY INFORMATION: The ‘‘Recognition of Additional Magisterial application to the Commissioner of provisions of 35 U.S.C. 156, ’’ Extension Districts as Citrus Black Spot Pest-Free Patents and Trademarks, Department of of patent term,’’ provide, generally, that Areas for the Republic of South Africa.’’ Commerce, for the extension of a patent a patent for a product may be extended The CIED may be viewed on the that claims that veterinary biologic. for a period of up to 5 years as long as Regulations.gov Web site or in our DATES: We will consider all requests for the patent claims a product that, among reading room (see ADDRESSES above for revision of the regulatory review period instructions for accessing other things, was subject to a regulatory determination that we receive on or review period before its commercial Regulations.gov and information on the before August 26, 2009. -

Review of Existing Infrastructure in the Orange River Catchment
Study Name: Orange River Integrated Water Resources Management Plan Report Title: Review of Existing Infrastructure in the Orange River Catchment Submitted By: WRP Consulting Engineers, Jeffares and Green, Sechaba Consulting, WCE Pty Ltd, Water Surveys Botswana (Pty) Ltd Authors: A Jeleni, H Mare Date of Issue: November 2007 Distribution: Botswana: DWA: 2 copies (Katai, Setloboko) Lesotho: Commissioner of Water: 2 copies (Ramosoeu, Nthathakane) Namibia: MAWRD: 2 copies (Amakali) South Africa: DWAF: 2 copies (Pyke, van Niekerk) GTZ: 2 copies (Vogel, Mpho) Reports: Review of Existing Infrastructure in the Orange River Catchment Review of Surface Hydrology in the Orange River Catchment Flood Management Evaluation of the Orange River Review of Groundwater Resources in the Orange River Catchment Environmental Considerations Pertaining to the Orange River Summary of Water Requirements from the Orange River Water Quality in the Orange River Demographic and Economic Activity in the four Orange Basin States Current Analytical Methods and Technical Capacity of the four Orange Basin States Institutional Structures in the four Orange Basin States Legislation and Legal Issues Surrounding the Orange River Catchment Summary Report TABLE OF CONTENTS 1 INTRODUCTION ..................................................................................................................... 6 1.1 General ......................................................................................................................... 6 1.2 Objective of the study ................................................................................................ -

Faith in an Anglo-Boer War Concentration Camp of 1901
Acta Academica 2007 39(3): 21-44 Dolf Britz ‘Now, what has become of our prayers and supplications?’ Faith in an Anglo-Boer War concentration camp of 1901 First submission: February 2007 Christian faith played a significant and decisive role in the concentration camps of the Anglo-Boer War (1899-1902). This article focuses on the diary of A D Luckhoff (1874-1963) who, after completing his theological training, volunteered for the ministry in the concentration camp at Bethulie. The anguish, distress and trials of this inex- perienced young clergyman are disclosed by allowing the diary to speak for itself, thus ensuring that its profound spiritual and emotional character is retained in the expo- sition. Alongside this exposition, observations and historical notes are appended, as well as questions concerning the diarist’s underlying theological convictions, which are compared and contrasted with those evinced in the contemporary diary of a young female resident of the camp, Rensche van der Walt (1878-1948). The ensuing analysis aims to demonstrate that Christian faith was a more complicated and complex issue during this war than has yet been realised. ‘Ag, wat het van ons gebede en versugtinge geword?’ Geloof in ’n konsentrasiekamp gedurende die Anglo- Boereoorlog 1901 Gedurende die Anglo-Boereoorlog (1899-1902) het die Christelike geloof ’n bepalende en belangrike rol gespeel in die konsentrasiekampe. Hierdie artikel gaan in op die dagboek van ’n jong aspirant predikant, A D Luckhoff (1874-1963). Hy het hom na voltooiing van sy teologiese opleiding beskikbaar gestel vir die bediening in die kon- sentrasiekamp van Bethulie. Sy dagboek word geanaliseer met die oog daarop om die spiritualiteit, onderliggende teologiese oortuigings, refleksie én aanvegting, bloot te lê. -

South African Jewish Board of Deputies Report of The
The South African Jewish Board of Deputies JL 1r REPORT of the Executive Council for the period July 1st, 1933, to April 30th, 1935. To be submitted to the Eleventh Congress at Johannesburg, May 19th and 20th, 1935. <י .H.W.V. 8. Co י É> S . 0 5 Americanist Commiitae LIBRARY 1 South African Jewish Board of Deputies. EXECUTIVE COUNCIL. President : Hirsch Hillman, Johannesburg. Vice-President» : S. Raphaely, Johannesburg. Morris Alexander, K.C., M.P., Cape Town. H. Moss-Morris, Durban. J. Philips, Bloemfontein. Hon. Treasurer: Dr. Max Greenberg. Members of Executive Council: B. Alexander. J. Alexander. J. H. Barnett. Harry Carter, M.P.C. Prof. Dr. S. Herbert Frankel. G. A. Friendly. Dr. H. Gluckman. J. Jackson. H. Katzenellenbogen. The Chief Rabbi, Prof. Dr. j. L. Landau, M.A., Ph.D. ^ C. Lyons. ^ H. H. Morris Esq., K.C. ^י. .B. L. Pencharz A. Schauder. ^ Dr. E. B. Woolff, M.P.C. V 2 CONSTITUENT BODIES. The Board's Constituent Bodies are as. follows :— JOHANNESBURG (Transvaal). 1. Anykster Sick Benefit and Benevolent Society. 2. Agoodas Achim Society. 3. Beth Hamedrash Hagodel. 4. Berea Hebrew Congregation. 5. Bertrams Hebrew Congregation. 6. Braamfontein Hebrew Congregation. 7. Chassidim Congregation. 8. Club of Polish Jews. 9. Doornfontein Hebrew:: Congregation.^;7 10. Eastern Hebrew Benevolent Society. 11. Fordsburg Hebrew Congregation. 12. Grodno Sifck Benefit and Benevolent Society. 13. Habonim. 14. Hatechiya Organisation. 15. H.O.D. Dr. Herzl Lodge. 16. H.O.D. Sir Moses Montefiore Lodge. 17. Jeppes Hebrew Congregation, 18. Johannesburg Jewish Guild. 19. Johannesburg Jewish Helping Hand and Burial Society. -

Arid Areas Report, Volume 1: District Socio�Economic Profile 2007 NO 1 and Development Plans
Arid Areas Report, Volume 1: District socio-economic profile 2007 NO 1 and development plans Arid Areas Report, Volume 1: District socio-economic profile and development plans Centre for Development Support (IB 100) University of the Free State PO Box 339 Bloemfontein 9300 South Africa www.ufs.ac.za/cds Please reference as: Centre for Development Support (CDS). 2007. Arid Areas Report, Volume 1: District socio-economic profile and development plans. CDS Research Report, Arid Areas, 2007(1). Bloemfontein: University of the Free State (UFS). CONTENTS I. Introduction ...................................................................................................................... 1 II. Geographic overview ........................................................................................................ 2 1. Namaqualand and Richtersveld ................................................................................................... 3 2. The Karoo................................................................................................................................... 4 3. Gordonia, the Kalahari and Bushmanland .................................................................................... 4 4. General characteristics of the arid areas ....................................................................................... 5 III. The Western Zone (Succulent Karoo) .............................................................................. 8 1. Namakwa District Municipality ..................................................................................................