The Selective Estrogen Enzyme Modulators in Breast Cancer: a Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

United States July 2016 2 Table of Contents
Deuterium Labelled Compounds United States July 2016 2 Table of Contents International Distributors 3 Corporate Overview 4 General Information 5 Pricing and Payment 5 Quotations 5 Custom Synthesis 5 Shipping 5 Quality Control 6 Quotations 6 Custom Synthesis 6 Shipping 6 Quality Control 6 Chemical Abstract Service Numbers 6 Handling Hazardous Compounds 6 Our Products are Not Intended for Use in Humans 7 Limited Warranty 7 Packaging Information 7 Alphabetical Listings 8 Stock Clearance 236 Products by Category 242 n-Alkanes 243 α-Amino Acids, N-Acyl α-Amino Acids, N-t-BOC Protected α-Amino Acid 243 and N-FMOC Protected α-Amino Acids Buffers and Reagents for NMR Studies 245 Detergents 245 Environmental Standards 246 Fatty Acids and Fatty Acid Esters 249 Flavours and Fragrances 250 Gases 253 Medical Research Products 254 Nucleic Acid Bases and Nucleosides 255 Pesticides and Pesticide Metabolites 256 Pharmaceutical Standards 257 Polyaromatic Hydrocarbons (PAHs), Alkyl-PAHs, Amino-PAHs, 260 Hydroxy-PAHs and Nitro-PAHs Polychlorinated Biphenyls (PCBs) 260 Spin Labels 261 Steroids 261 3 International Distributors C Beijng Zhenxiang H EQ Laboratories GmbH Australia K Technology Company Graf-von-Seyssel-Str. 10 Rm. 15A01, Changyin Bld. 86199 Augsburg Austria H No. 88, YongDingLu Rd. Germany Beijing 100039 Tel.: (49) 821 71058246 Belgium J China Fax: (49) 821 71058247 Tel.: (86) 10-58896805 [email protected] China C Fax: (86) 10-58896158 www.eqlabs.de Czech Republic H [email protected] Germany, Austria, China Czech Republic, Greece, Denmark I Hungary, -
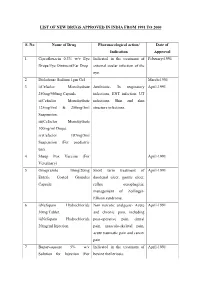
List of New Drugs Approved in India from 1991 to 2000
LIST OF NEW DRUGS APPROVED IN INDIA FROM 1991 TO 2000 S. No Name of Drug Pharmacological action/ Date of Indication Approval 1 Ciprofloxacin 0.3% w/v Eye Indicated in the treatment of February-1991 Drops/Eye Ointment/Ear Drop external ocular infection of the eye. 2 Diclofenac Sodium 1gm Gel March-1991 3 i)Cefaclor Monohydrate Antibiotic- In respiratory April-1991 250mg/500mg Capsule. infections, ENT infection, UT ii)Cefaclor Monohydrate infections, Skin and skin 125mg/5ml & 250mg/5ml structure infections. Suspension. iii)Cefaclor Monohydrate 100mg/ml Drops. iv)Cefaclor 187mg/5ml Suspension (For paediatric use). 4 Sheep Pox Vaccine (For April-1991 Veterinary) 5 Omeprazole 10mg/20mg Short term treatment of April-1991 Enteric Coated Granules duodenal ulcer, gastric ulcer, Capsule reflux oesophagitis, management of Zollinger- Ellison syndrome. 6 i)Nefopam Hydrochloride Non narcotic analgesic- Acute April-1991 30mg Tablet. and chronic pain, including ii)Nefopam Hydrochloride post-operative pain, dental 20mg/ml Injection. pain, musculo-skeletal pain, acute traumatic pain and cancer pain. 7 Buparvaquone 5% w/v Indicated in the treatment of April-1991 Solution for Injection (For bovine theileriosis. Veterinary) 8 i)Kitotifen Fumerate 1mg Anti asthmatic drug- Indicated May-1991 Tablet in prophylactic treatment of ii)Kitotifen Fumerate Syrup bronchial asthma, symptomatic iii)Ketotifen Fumerate Nasal improvement of allergic Drops conditions including rhinitis and conjunctivitis. 9 i)Pefloxacin Mesylate Antibacterial- In the treatment May-1991 Dihydrate 400mg Film Coated of severe infection in adults Tablet caused by sensitive ii)Pefloxacin Mesylate microorganism (gram -ve Dihydrate 400mg/5ml Injection pathogens and staphylococci). iii)Pefloxacin Mesylate Dihydrate 400mg I.V Bottles of 100ml/200ml 10 Ofloxacin 100mg/50ml & Indicated in RTI, UTI, May-1991 200mg/100ml vial Infusion gynaecological infection, skin/soft lesion infection. -

C:\Data\Ndaenjuvia\AP LTR 05-07-04
NDA 21-443 Package Insert ENJUVIA™ (synthetic conjugated estrogens, B) Tablets Rx only ESTROGENS INCREASE THE RISK OF ENDOMETRIAL CANCER Close clinical surveillance of all women taking estrogens is important. Adequate diagnostic measures, including endometrial sampling when indicated, should be undertaken to rule out malignancy in all cases of undiagnosed persistent or recurring abnormal vaginal bleeding. There is no evidence that the use of “natural” estrogens results in a different endometrial risk profile than synthetic estrogens at equivalent estrogen doses. (See WARNINGS, Malignant neoplasms, Endometrial cancer.) CARDIOVASCULAR AND OTHER RISKS Estrogens with or without progestins should not be used for the prevention of cardiovascular disease. (See WARNINGS, Cardiovascular disorders.) The Women’s Health Initiative (WHI) study reported increased risks of myocardial infarction, stroke, invasive breast cancer, pulmonary emboli, and deep vein thrombosis in postmenopausal women (50 to 79 years of age) during 5 years of treatment with oral conjugated estrogens (CE 0.625 mg) combined with medroxyprogesterone acetate (MPA 2.5 mg) relative to placebo. (See CLINICAL PHARMACOLOGY, Clinical Studies). The Women’s Health Initiative Memory Study (WHIMS), a substudy of WHI, reported increased risk of developing probable dementia in postmenopausal women 65 years of age or older during 4 years of treatment with oral conjugated estrogens plus medroxyprogesterone acetate relative to placebo. It is unknown whether this finding applies to younger postmenopausal women or to women taking estrogen alone therapy. (See CLINICAL PHARMACOLOGY, Clinical Studies.) Other doses of oral conjugated estrogens with medroxyprogesterone acetate, and other combinations and dosage forms of estrogens and progestins were not studied in the WHI clinical trials and, in the absence of comparable data, these risks should be assumed to be similar. -

Selective Spatial Upregulation of Intratumoral Stromal Aromatase in Breast Cancer Patients: Evidence for Imbalance of Local Estrogen Metabolism
Endocrine-Related Cancer (2006) 13 1101–1107 Selective spatial upregulation of intratumoral stromal aromatase in breast cancer patients: evidence for imbalance of local estrogen metabolism Christian F Singer1,2, Anneliese Fink-Retter1, Daphne Gschwantler-Kaulich1, Theresia Thalhammer 3, Gernot Hudelist1, Ruth Mueller1,2, Klaus Czerwenka4 and Ernst Kubista1,2 1Division of Special Gynecology, University of Vienna Medical Center, Waehringer Guertel 18-20, 1090, Vienna, Austria 2Ludwig-Boltzmann-Institute of Clinical Experimental Oncology, 3Center for Physiology and Pathophysiology, 4Division of Gynecopathology, Medical University of Vienna, Vienna, Austria (Requests for offprints should be addressed to C F Singer; Email: [email protected]) Abstract The suppression of local estrogens levels is of key importance in the treatment of ER-positive breast cancer. Essentially all endocrine strategies act by either suppressing estrogen formation or competitively inhibiting receptor-binding in tumor cells. Nevertheless, little is still known about the local expression of aromatase and sulfotransferase which are the key modulators of intra-tumoral estrogen levels. We have performed immunohistochemostry to investigate the expression of aromatase and sulfotransferase in 42 samples obtained directly from malignant breast tumors, and compared it to biopsies obtained from uninvolved tissue in the vicinity of the invasion front, and to distant breast tissue. We found that aromatase was equally detectable in both tumor epithelial and stroma, but was mostly epithelial in non-malignant tissues (PZ0.00008, Fisher’s exact test). Also, aromatase protein expression was significantly more common in tumoral stroma when compared with peritumoral and distant breast stroma (PZ0.00005, and P!0.00001 respectively). With the notable exception of cystosarcoma phylloides, sulfotransferase protein was detectable only in epithelial tissues, regardless of the location within the diseased breast. -

Classification and Pharmacology of Progestins
Maturitas 46S1 (2003) S7–S16 Classification and pharmacology of progestins Adolf E. Schindler a,∗, Carlo Campagnoli b, René Druckmann c, Johannes Huber d, Jorge R. Pasqualini e, Karl W. Schweppe f, Jos H. H. Thijssen g a Institut für Medizinische Forschung und Fortbildung, Universitätsklinikum, Hufelandstr. 55, Essen 45147, Germany b Ospedale Ginecologico St. Anna, Corso Spezia 60, 10126 Torino, Italy c Ameno-Menopause-Center, 12, Rue de France, 06000 Nice, France d Abt. für Gynäkologische Endokrinologie, AKH Wien, Währingergürtel 18-20, 1090 Wien, Austria e Institute de Puériculture26, Boulevard Brune, 75014 Paris, France f Abt. für Gynäkologie und Geburtshilfe, Ammerland Klinik, Langestr.38, 26622 Westerstede, Germany g Department of Endocrinology, Universitair Medisch Centrum Utrecht, P.O. Box 85090, 3508 AB Utrecht, The Netherlands Abstract Besides the natural progestin, progesterone, there are different classes of progestins, such as retroprogesterone (i.e. dydroges- terone), progesterone derivatives (i.e. medrogestone) 17␣-hydroxyprogesterone derivatives (i.e. chlormadinone acetate, cypro- terone acetate, medroxyprogesterone acetate, megestrol acetate), 19-norprogesterone derivatives (i.e. nomegestrol, promege- stone, trimegestone, nesterone), 19-nortestosterone derivatives norethisterone (NET), lynestrenol, levonorgestrel, desogestrel, gestodene, norgestimate, dienogest) and spironolactone derivatives (i.e. drospirenone). Some of the synthetic progestins are prodrugs, which need to be metabolized to become active compounds. Besides -

Northern Ireland Prescription Code Book Drugs Section September 2021
Family Practitioner Services, Pharmaceutical Services, 2 Franklin Street, Belfast BT2 8DQ Telephone No. 028 9053 5613, Fax No. 028 9053 2963 Northern Ireland Prescription Code Book Drugs section September 2021 Page 1 of 587 Effective for prescriptions dispensed September 2021 BSO Code dm+d Pack Description Code Number Special ZD Of Container 38960 4SURE beta-ketone testing strips 4S-810-4183401-001 (Nipro Diagnostics (UK) Ltd) 10 strip strips 38961 4SURE testing strips (Nipro Diagnostics (UK) Ltd) 50 strip strips 4144 AAA 1.5mg/dose sore throat spray (Manx Healthcare Ltd) 60 dose [BSO pack = 1] BSO Unit Of Measure Code No. devices dispensed 70598 (DT) Abacavir 600mg / Lamivudine 300mg tablets 30 tablet tablets 9154 Abasaglar 100units/ml solution for injection 3ml cartridges (Eli Lilly and Company Ltd) 5 cartridge cartridges ZD 9038 Abasaglar KwikPen 100units/ml solution for injection 3ml pre-filled pens (Eli Lilly and Company Ltd) 5 pre- injections ZD filled disposable injection 70088 (DT) Abatacept 125mg/1ml solution for injection pre-filled disposable devices 4 pre-filled disposable injections ZD injection 70089 (DT) Abatacept 125mg/1ml solution for injection pre-filled syringes 4 pre-filled disposable injection injections ZD 70144 Abelcet 100mg/20ml concentrate for suspension for infusion vials (Teva UK Ltd) 10 vial vials Hospital Only 3303 Abidec Multivitamin drops (Omega Pharma Ltd) 25 ml mls 4043 Abilify 10mg orodispersible tablets (Otsuka Pharmaceuticals (U.K.) Ltd) 28 tablet 4 x 7 tablets tablets 3006 Abilify 10mg tablets (Otsuka -

209627Orig1s000
CENTER FOR DRUG EVALUATION AND RESEARCH APPLICATION NUMBER: 209627Orig1s000 MULTI-DISCIPLINE REVIEW Summary Review Office Director Cross Discipline Team Leader Review Clinical Review Non-Clinical Review Statistical Review Clinical Pharmacology Review Reviewers of Multi-Disciplinary Review and Evaluation SECTIONS OFFICE/ AUTHORED/ ACKNOWLEDGED/ DISCIPLINE REVIEWER DIVISION APPROVED Mark Seggel, Ph.D. OPQ/ONDP/DNDP2 Authored: Section 4.2 Digitally signed by Mark R. Seggel -S CMC Lead DN: c=US, o=U.S. Government, ou=HHS, ou=FDA, ou=People, cn=Mark R. Signature: Mark R. Seggel -S Seggel -S, 0.9.2342.19200300.100.1.1=1300071539 Date: 2018.08.08 16:29:15 -04'00' Frederic Moulin, DVM, PhD OND/ODE3/DBRUP Authored: Section 5 Pharmacology/ Digitally signed by Frederic Moulin -S Toxicology DN: c=US, o=U.S. Government, ou=HHS, ou=FDA, ou=People, Reviewer Signature: Frederic Moulin -S 0.9.2342.19200300.100.1.1=2001708658, cn=Frederic Moulin -S Date: 2018.08.08 15:26:57 -04'00' Kimberly Hatfield, PhD OND/ODE3/DBRUP Approved: Section 5 Pharmacology/ Toxicology Digitally signed by Kimberly P. Hatfield -S DN: c=US, o=U.S. Government, ou=HHS, ou=FDA, ou=People, Team Leader Signature: Kimberly P. Hatfield -S 0.9.2342.19200300.100.1.1=1300387215, cn=Kimberly P. Hatfield -S Date: 2018.08.08 14:56:10 -04'00' Li Li, Ph.D. OCP/DCP3 Authored: Sections 6 and 17.3 Clinical Pharmacology Dig ta ly signed by Li Li S DN c=US o=U S Government ou=HHS ou=FDA ou=People Reviewer cn=Li Li S Signature: Li Li -S 0 9 2342 19200300 100 1 1=20005 08577 Date 2018 08 08 15 39 23 04'00' Doanh Tran, Ph.D. -

A Review of Mastalgia in Patients with Fibrocystic Breast Changes and the Non-Surgical Treatment Options
REVIEW ARTICLE A Review of Mastalgia in Patients with Fibrocystic Breast Changes and the Non-Surgical Treatment Options Khalid Rida Murshid FRCS(C), FACS Department of Surgery, College of Medicine, Taibah University, Al Madinah Al Munawwarah, Kingdom of Saudi Arabia Abstract Objectives The objectives of this study are to review fibrocystic changes of the breast, their causal and associated factors and their correlation to mastalgia, and then to review the available treatment options for mastalgia (caused by fibrocystic changes) short of surgery. Methods The author reviews all the articles obtained from a PubMed research on mastalgia and fibrocystic changes of the breast, published in English over the last 14 years. Results Fibrocystic changes of the breast are common and can be considered as a normal phase of breast development. These changes are sometimes asymptomatic; however, when painful, patients would seek medical advice. Lifestyle changes and the avoidance of certain dietary elements as well as the use of some non-pharmacological agents have shown some beneficiary effects. In severe cases, stronger pharmacological and hormonal agents are resorted to being more effective but are associated with greater side effects. Conclusion Fibrocystic changes of the breast are common and should not be considered a disease. When painful, reassurance and non-pharmacological measures should be used first as a treatment. Stronger pharmacological and hormonal agents hold more serious side effects. Some of these remedies are supported by good clinical evidence, while others are not. The ideal treatment for mastalgia caused by fibrocystic changes is to be identified by sound recent randomized controlled clinical studies on simple remedies before being performed on stronger ones. -

Vaginal Delivery System
(19) & (11) EP 2 062 568 A1 (12) EUROPEAN PATENT APPLICATION (43) Date of publication: (51) Int Cl.: 27.05.2009 Bulletin 2009/22 A61K 9/00 (2006.01) (21) Application number: 07397042.8 (22) Date of filing: 22.11.2007 (84) Designated Contracting States: • Hanes, Vladimir AT BE BG CH CY CZ DE DK EE ES FI FR GB GR Tarrytown, NY 10591 (US) HU IE IS IT LI LT LU LV MC MT NL PL PT RO SE • Keinänen, Antti SI SK TR 20540 Turku (FI) Designated Extension States: • Holmberg, Svante AL BA HR MK RS 20900 Turku (FI) • Nikander, Hannu (71) Applicant: Bayer Schering Pharma Oy 21330 Paattinen (FI) 20210 Turku (FI) (74) Representative: Matilainen, Mirja Helena et al (72) Inventors: Oy Jalo Ant-Wuorinen AB, • Talling, Christine Iso Roobertinkatu 4-6 A 20610 Turku (FI) 00120 Helsinki (FI) (54) Vaginal delivery system (57) The present invention is related to an intravag- brane (3) encasing the core, said core and membrane inal delivery system for the controlled release of a pro- essentially consisting of a same or different polymer com- gestogen and an estrogen, comprising additionally a position, wherein at cast one of the cores comprises a therapeutically active or a health-promoting substance progestogen or a mixture of a progestogen and an es- (1) capable of giving and/or enhancing the protection trogen, and another core may comprise an estrogen or against bacterial and fungal infections, and/or enhancing a progestogen, and wherein the membrane or the surface the protection against sexually transmitted diseases. The of the membrane or at least one of the cores comprises delivery system consists of one or more compartments said therapeutically active or a health-promoting sub- (2,4,5), one of each comprising a core (7) and a mem- stance. -

The Importance of Thyroid Hormone Sulfation During Fetal Development
The importance of thyroid hormone sulfation during fetal development Monique H.A. Kester CIP-data Koninklijke Bibliotheek, Den Haag Kester, Monique Helene Agathe The importance of thyroid hormone sulfation during fetal development Thesis Erasmus University Rotterdam - with summary in Dutch ISBN 90 ·901501 0-2 Cover: White Rabbit Photo I Gert-Jan van den Bemd ~ Printed by Ridderprint offsetdrukkerij BV, Ridderkerk The research described in this thesis was financially supported by the Sophia Foundation for Medical Research (project 211). The publication of this thesis was financially supported by Apotheek J.H. van Waert BV and Organon Nederland BV. The importance of thyroid hormone sulfation during fetal development Het belang van schildklierhormoonsulfatering tijdens de foetale ontwikkeling Proefschrift ter verkrijging van de graad van doctor aan de Erasmus Universiteit Rotterdam op gezag van de Rector Magnificus Prof.dr.ir. J.H. van Semmel en volgens besluit van het College voor Promoties De openbare verdediging zal plaatsvinden op woensdag 5 september 2001 om 15.45 uur door Monique Helene Agathe Kester geboren te Sint-Michielsgestel Promotiecommisie Promotoren: Prof.dr.ir. T.J. Visser Prof.dr. D. Tibboel Overige leden: Prof.dr. F.H. de Jong Prof.dr. A. Brouwer Dr. M.W.H. Coughtrie The research described in this thesis was performed at the Departments of Internal Medicine and Pediatric Surgery of the Erasmus University Medical Center Rotterdam, The Netherlands. Labor improbius omnia vinci! De aanhouder win! Vergilius Contents List of abbreviations -

Etats Rapides
List of European Pharmacopoeia Reference Standards Effective from 2015/12/24 Order Reference Standard Batch n° Quantity Sale Information Monograph Leaflet Storage Price Code per vial Unit Y0001756 Exemestane for system suitability 1 10 mg 1 2766 Yes +5°C ± 3°C 79 ! Y0001561 Abacavir sulfate 1 20 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0001552 Abacavir for peak identification 1 10 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0001551 Abacavir for system suitability 1 10 mg 1 2589 Yes +5°C ± 3°C 79 ! Y0000055 Acamprosate calcium - reference spectrum 1 n/a 1 1585 79 ! Y0000116 Acamprosate impurity A 1 50 mg 1 3-aminopropane-1-sulphonic acid 1585 Yes +5°C ± 3°C 79 ! Y0000500 Acarbose 3 100 mg 1 See leaflet ; Batch 2 is valid until 31 August 2015 2089 Yes +5°C ± 3°C 79 ! Y0000354 Acarbose for identification 1 10 mg 1 2089 Yes +5°C ± 3°C 79 ! Y0000427 Acarbose for peak identification 3 20 mg 1 Batch 2 is valid until 31 January 2015 2089 Yes +5°C ± 3°C 79 ! A0040000 Acebutolol hydrochloride 1 50 mg 1 0871 Yes +5°C ± 3°C 79 ! Y0000359 Acebutolol impurity B 2 10 mg 1 -[3-acetyl-4-[(2RS)-2-hydroxy-3-[(1-methylethyl)amino] propoxy]phenyl] 0871 Yes +5°C ± 3°C 79 ! acetamide (diacetolol) Y0000127 Acebutolol impurity C 1 20 mg 1 N-(3-acetyl-4-hydroxyphenyl)butanamide 0871 Yes +5°C ± 3°C 79 ! Y0000128 Acebutolol impurity I 2 0.004 mg 1 N-[3-acetyl-4-[(2RS)-3-(ethylamino)-2-hydroxypropoxy]phenyl] 0871 Yes +5°C ± 3°C 79 ! butanamide Y0000056 Aceclofenac - reference spectrum 1 n/a 1 1281 79 ! Y0000085 Aceclofenac impurity F 2 15 mg 1 benzyl[[[2-[(2,6-dichlorophenyl)amino]phenyl]acetyl]oxy]acetate -

Giving Another Chance to Mifepristone in Pharmacotherapy for Aggressive Meningiomas—A Likely Synergism with Hydroxyurea?
Curr Probl Cancer 40 (2016) 229–243 Contents lists available at ScienceDirect Curr Probl Cancer journal homepage: www.elsevier.com/locate/cpcancer Giving another chance to mifepristone in pharmacotherapy for aggressive meningiomas—A likely synergism with hydroxyurea? İlhan Elmaci, MDa, Meric A. Altinoz, MDb,*, Aydin Sav, MDc, Zeliha Yazici, PhDd, Aysel Ozpinar, PhDe Introduction Meningiomas: An underestimated health problem Meningiomas are the most frequently reported intracranial tumors, accounting for approx- imately one-fourth of all reported primary brain neoplasms.1 They are benign in approximately 90% of the cases and the remaining cases are either borderline or atypical (World Health Organization [WHO] grade II) or malignant (WHO grade III).1 In the United States, the incidence rates with similar age standardization estimated from figures provided by the Central Brain Tumor Registrywere1.8formenand4.2per100,000forwomenin2006.1 Increasing incidence rates of meningiomas have been reported from several industrialized countries since the early 1980s. But recent studies revealed that real meningioma incidence is even higher.1 As meningiomas are mostly benign, they are not covered by most cancer registries.1 Nevertheless, in Finland, as in other Nordic countries, all surgeons and pathologists are obliged to report all tumors of the central nervous system, both malignant and benign, to the cancer registry.1 To reveal the real incidence of Grant/Financial Support: none. This is original work and is not under consideration for publication elsewhere. This submission is for the special issue being compiled by Beata Holkova on the topic Amylodosis. a Department of Neurosurgery, Memorial Hospital, Istanbul, Turkey b Department of Immunology, Experimental Medical Research Institute/DETAE, Istanbul University, Istanbul, Turkey c Nisantasi Neuropathology Group, Istanbul, Turkey d Department of Pharmacology, Cerrahpasa Faculty of Medicine, Istanbul University, Turkey e Department of Biochemistry, Acibadem University, Istanbul, Turkey * Corresponding author: Meric A.