Population Energetics of the American White Ibis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

156 Glossy Ibis
Text and images extracted from Marchant, S. & Higgins, P.J. (co-ordinating editors) 1990. Handbook of Australian, New Zealand & Antarctic Birds. Volume 1, Ratites to ducks; Part B, Australian pelican to ducks. Melbourne, Oxford University Press. Pages 953, 1071-1 078; plate 78. Reproduced with the permission of Bird life Australia and Jeff Davies. 953 Order CICONIIFORMES Medium-sized to huge, long-legged wading birds with well developed hallux or hind toe, and large bill. Variations in shape of bill used for recognition of sub-families. Despite long legs, walk rather than run and escape by flying. Five families of which three (Ardeidae, Ciconiidae, Threskiornithidae) represented in our region; others - Balaenicipitidae (Shoe-billed Stork) and Scopidae (Hammerhead) - monotypic and exclusively Ethiopian. Re lated to Phoenicopteriformes, which sometimes considered as belonging to same order, and, more distantly, to Anseriformes. Behavioural similarities suggest affinities also to Pelecaniformes (van Tets 1965; Meyerriecks 1966), but close relationship not supported by studies of egg-white proteins (Sibley & Ahlquist 1972). Suggested also, mainly on osteological and other anatomical characters, that Ardeidae should be placed in separate order from Ciconiidae and that Cathartidae (New World vultures) should be placed in same order as latter (Ligon 1967). REFERENCES Ligon, J.D. 1967. Occas. Pap. Mus. Zool. Univ. Mich. 651. Sibley, C. G., & J.E. Ahlquist. 1972. Bull. Peabody Mus. nat. Meyerriecks, A.J. 1966. Auk 83: 683-4. Hist. 39. van Tets, G.F. 1965. AOU orn. Monogr. 2. 1071 Family PLATALEIDAE ibises, spoonbills Medium-sized to large wading and terrestial birds. About 30 species in about 15 genera, divided into two sub families: ibises (Threskiornithinae) and spoonbills (Plataleinae); five species in three genera breeding in our region. -

IUCN-SSC Stork, Ibis and Spoonbill Specialist Group Special Publication 2
IUCN-SSC Stork, Ibis and Spoonbill Specialist Group Special Publication 2 Proceedings of the IX Workshop of the AEWA Eurasian Spoonbill International Expert Group Djerba Island, Tunisia, 14th - 18th November 2018 Editors: Jocelyn Champagnon, Jelena Kralj, Luis Santiago Cano Alonso and K. S. Gopi Sundar Editors-in-Chief, Special Publications, IUCN-SSC Stork, Ibis and Spoonbill Specialist Group K.S. Gopi Sundar, Co-chair IUCN Stork, Ibis and Spoonbill Specialist Group Luis Santiago Cano Alonso, Co-chair IUCN Stork, Ibis and Spoonbill Specialist Group Invited Editors for this issue Jocelyn Champagnon, Tour du Valat, Research Institute for the Conservation of Mediterranean Wetlands, Arles, France Jelena Kralj, Institute of Ornithology, Zagreb, Croatia Expert Review Board Hichem Azafzaf, Association “les Amis des Oiseaux » (AAO/BirdLife Tunisia), Tunisia Petra de Goeij, Royal NIOZ, the Netherlands Csaba Pigniczki, Kiskunság National Park Directorate, Hungary Suggested citation of this publication: Champagnon J., Kralj J., Cano Alonso, L. S. & Sundar, K. S. G. (ed.) 2019. Proceedings of the IX Workshop of the AEWA Eurasian Spoonbill International Expert Group. IUCN-SSC Stork, Ibis and Spoonbill Specialist Group Special Publication 2. Arles, France. ISBN 978-2-491451-00-4. Recommended Citation of a chapter: Marion L. 2019. Recent trends of the breeding population of Spoonbill in France 2012- 2018. Pp 19- 23. In: Champagnon J., Kralj J., Cano Alonso, L. S. & Sundar, K. S. G. (ed.) Proceedings of the IX Workshop of the AEWA Eurasian Spoonbill International Expert Group. IUCN-SSC Stork, Ibis and Spoonbill Specialist Group Special Publication 2. Arles, France. INFORMATION AND WRITING DISCLAIMER The information and opinions expressed in this publication belong to the authors. -
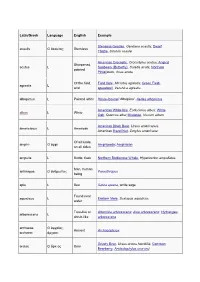
Dwarf Thistle, Cirsi
Latin/Greek Language English Example Stemless Gentian, Gentiana acaulis; Dwarf acaulis G ἄκαυλος Stemless Thistle, Cirsium acaule American Crocodile, Crocodylus acutus; Angled Sharpened, acutus L Sunbeam (Butterfly), Curetis acuta; Northern pointed Pintailduck, Anas acuta Of the field, Field Vole, Microtus agrestis; Green Field- agrestis L wild speedwell, Veronica agrestis albopictus L Painted white Hosta fortunei 'Albopicta', Aedes albopictus American White Ibis, Eudocimus albus; White albus L White Oak, Quercus alba; Mistletoe, Viscum album American Black Bear, Ursus americanus; americanus L American American Hazel Nut, Corylus americana Of all kinds, amphi- G ἀμφί Amphipoda; Amphibian on all sides ampulla L Bottle, flask Northern Bottlenose Whale, Hyperoodon ampullatus Man, human anthropos G ἄνθρωπος Paranthropus being apis L Bee Salvia apiana, white sage Found near aquaticus L Eastern Mole, Scalopus aquaticus water Tree-like or Artemisia arborescens; Aloe arborescens; Hydrangea arborescens L shrub-like arborescens archaeos, G ἀρχαῖος, Ancient Archaeopteryx archaeo- ἀρχαιο- Grizzly Bear, Ursus arctos horribilis; Common arctos G ἄρκτος Bear Bearberry, Arctostaphylos uva-ursi argentatus L Silvery Herring Gull, Larus argentatus arthron G ἄρθρον Joint Arthropoda arvensis L In the field Skylark, Alauda arvensis astron, astro-, G ἄστρον, Star Starfish (class), Asteroidea astero- ἀστρο-, ἀστερο- Acer palmatum 'Atropurpureum'; Berberis atropurpureum L Deep purple thunbergii f. atropurpurea Daphne odora 'Aureomarginata'; Taxus aureomarginata -

Flocking of V-Shaped and Echelon Northern Bald Ibises with Different Wingspans: Repositioning and Energy Saving
Flocking of V-shaped and Echelon Northern Bald Ibises with Different Wingspans: Repositioning and Energy Saving Amir Mirzaeinia1, Mehdi Mirzaeinia2, Mostafa Hassanalian3* V-shaped and echelon formations help migratory birds to consume less energy for migration. As the case study, the formation flight of the Northern Bald Ibises is considered to investigate different effects on their flight efficiency. The effects of the wingtip spacing and wingspan are examined on the individual drag of each Ibis in the flock. Two scenarios are considered in this study, (1) increasing and (2) decreasing wingspans toward the tail. An algorithm is applied for replacement mechanism and load balancing of the Ibises during their flight. In this replacing mechanism, the Ibises with the highest value of remained energy are replaced with the Ibises with the lowest energy, iteratively. The results indicate that depending on the positions of the birds with various sizes in the flock, they consume a different level of energy. Moreover, it is found that also small birds have the chance to take the lead during the flock. I. Introduction There are 1800 species of birds that migrate through seasonal long-distance routes from hundreds up to several thousand kilometers1. For example, Bar-tailed godwits as one of the non-stop migratory birds travel for 11600 km1,2. Canada Geese with an average speed of about 18 m/s migrate for 3,200 to 4,800 Km in a single day if the weather is good44. The Arctic terns migrate from the Arctic to the Antarctic1. Besides the mentioned migratory birds, other bird species including pelicans, swans, cormorants, cranes, and Ibis migrate in an echelon or V-shaped formations1. -

Developing a Research Network on Glossy Ibis, a Neglected Cosmopolitan Species
Developing a research network on Glossy ibis, a neglected cosmopolitan species Álvaro Arenas Patiño © 2014 The Project in a nutshell The Glossy ibis (Plegadis falcinellus) is the only cosmopolitan species of the Threskiornithidae family (ibis and spoonbills) and among the most widely distributed bird species in the world ([1] see map at the bottom of the document). Nonetheless, no knowledge exists on the intraspecific phylogeny and very little is known about the (meta)populations dynamics. We aim to fill this gap by setting up a research network and by implementing modern analytical tools (e.g. molecular and statistical) we nowadays dispose of. Some puzzling aspects of Glossy ibis abundance and distribution Little is known about the movement ecology of this species especially when focusing on a large spatial scale. In most ornithology books the Glossy ibis is described as migratory with nomadic elements [2] with populations at tropics being more sedentary than others. However, references are rarely provided and, to the best of our knowledge, only a few data have been ever collected on the Glossy ibis dispersal behavior. According to historical records, breeding was very rare in W Europe while common in North Africa and S Spain before the 20th century when it became almost absent in all these regions [3–5]. In 1996 seven pairs settled at Doñana, a wetland area in SW Spain, from where the species begun an astonishing population increase reaching about 8,000 pairs in current times [6,7]. Since then, the Glossy ibis has started to colonize other areas in W Europe and the Mediterranean Basin from where it had been absent for decades or even had never been recorded as a breeder species. -

New Observations of the Andean Ibis (Theristicus Branickii
SIT Graduate Institute/SIT Study Abroad SIT Digital Collections Independent Study Project (ISP) Collection SIT Study Abroad Fall 12-1-2014 New Observations of the Andean Ibis (Theristicus branickii, Threskiornithidae): Distribution, Movements, and Behavior Near Volcán Antisana Benjamin West SIT Study Abroad Follow this and additional works at: https://digitalcollections.sit.edu/isp_collection Part of the Latin American Studies Commons, Other Ecology and Evolutionary Biology Commons, Population Biology Commons, and the Zoology Commons Recommended Citation West, Benjamin, "New Observations of the Andean Ibis (Theristicus branickii, Threskiornithidae): Distribution, Movements, and Behavior Near Volcán Antisana" (2014). Independent Study Project (ISP) Collection. 2019. https://digitalcollections.sit.edu/isp_collection/2019 This Article is brought to you for free and open access by the SIT Study Abroad at SIT Digital Collections. It has been accepted for inclusion in Independent Study Project (ISP) Collection by an authorized administrator of SIT Digital Collections. For more information, please contact [email protected]. New Observations of the Andean Ibis (Theristicus branickii , Threskiornithidae): Distribution, Movements, and Behavior Near Volcán Antisana West, Benjamin M. Academic Directors: Silva, Xavier and Robayo, Javier Project Advisor: Williamson, Jessie Bowdoin College Biology South America, Ecuador, Napo Province, Reserva Ecológica Antisana Submitted in partial fulfillment of the requirements for Ecuador: Comparative Ecology and Conservation, SIT Study Abroad, Fall 2014 SIT Ecuador: Ecology, Fall 2014 West Abstract The Andean Ibis (Theristicus branickii ) of the highland grasslands of Ecuador, Peru, and Bolivia is listed globally as Near Threatened and Critically Endangered in Ecuador. The Ecuadorian population is estimated at 100 individuals and is restricted to the vicinities of Volcán Antisana and Volcán Cotopaxi. -

The Bare Head of the Northern Bald Ibis (Geronticus Eremita) Fulfills a Thermoregulatory Function Ismael Galván1* , Daniel Palacios2 and Juan José Negro1
Galván et al. Frontiers in Zoology (2017) 14:15 DOI 10.1186/s12983-017-0201-5 RESEARCH Open Access The bare head of the Northern bald ibis (Geronticus eremita) fulfills a thermoregulatory function Ismael Galván1* , Daniel Palacios2 and Juan José Negro1 Abstract Background: Dark pigments provide animals with several adaptive benefits such as protection against ultraviolet (UV) radiation and mechanical abrasion, but may also impose several constraints like a high absorbance of solar radiation. Endotherms, with relatively constant and high body temperatures, may be especially prone to thermoregulatory limitations if dark coloured and inhabiting hot environments. It is therefore expected that adaptations have specifically evolved because of these limitations. Bare, highly vascularised head skin may have evolved in birds with dark plumage from hot geographical regions because of favouring heat dissipation. Using the Northern bald ibis (Geronticus eremita) as a model species, we measured the surface temperature (Tsurf) of the head, the bill and the black feathered body of 11 birds along ambient temperatures (Ta) ranging from 21 to 42.5 °C employing thermal imaging. Results: While Tsurf of the bill and the feathered body was only slightly above Ta, head Tsurf was considerably higher, by up to 12 °C. Estimated values of heat loss followed similar variations. We also found that the red colour intensity of the head of ibises increased with head Tsurf, suggesting that birds are capable of controlling blood flow and the thermoregulatory function of the head. Conclusions: These findings are consistent with the hypothesis that bare skin has evolved in dark pigmented birds inhabiting hot environments because of their ability to dissipate heat. -

Plumage and Behavioral Development of Nestling White Ibises
Wilson Bull., 102(2), 1990, pp. 226-238 PLUMAGE AND BEHAVIORAL DEVELOPMENT OF NESTLING WHITE IBISES TONI L. DE SANTO,’ SUSAN G. MCDOWELL,~ AND KEITH L. BILDSTEIN~ Ans-rticr.- We describe the physical characteristicsand behavioral development OI 17 hand-reared and more than 400 parent-reared nestling White Ibises (Eudocimus albus) hatched in 1985 through 1988 at Pumpkinseed Island, a large colony site in coastal South Carolina. Hatchling ibises are covered with a Pale Neutral Gray to Jet Black natal plumage. About 30% of the hatchlingspossess a tuft of white feathers on their crown, and this pattern persiststhroughout the nestling period. Juvenal plumage, which is complete by 60 days, is mainly Vandyke Brown and Blackish Neutral Gray dorsally and creamy white ventrally. The bill, which is straight at hatching, begins to curve downward at about 14 days. Nestling White Ibises exhibit considerableindividual variation in bill markings from approximately 10 days of age through fledging. Increasingly persistent beggingvocalizations begin within hours of hatching. Nestlings walk on partially extended legsat eight days of age, pirate food from other nestlingsand form crechesat 2 1 days of age, and fledge and join all juvenile and mixed-age feeding flocks at 45-55 days of age. We suggestthat the phenotypic variability in plumage, bill coloration, and beggingcalls we describeenables parental ibises to identify more easily their offspringat the colony site. Received27 Feb. 1989, accepted12 Nov. 1989. Although the plumage and behavioral development of several species of wading birds has been studied in considerable detail (e.g., Hammerkops [&opus umbretta], Wilson et al. 1988; storks, Kahl 1962, 1966; Thomas 1984; herons, Gross 1923; Gavin0 and Dickerman 1972; Juarez and Dickerman 1972; Mc Vaugh 1972,1975; Snow 1974; Merritt 198 l), there are few detailed studies of juvenile ibises. -

THE BRITISH LIST the Official List of Bird Species Recorded in Britain Sponsored by Leica Sports Optics
THE BRITISH LIST The official list of bird species recorded in Britain sponsored by Leica Sports Optics CATEGORY E SPECIES Category E comprises those species that have been recorded as introductions, human-assisted transportees or escapees from captivity, whose breeding populations (if any) are thought not to be self-sustaining. Species in Category E that have bred in the wild in Britain are designated as E*. Species listed in Category E form no part of the British List, unless they are also included in Categories A, B or C. Descriptions and definitions of Categories of the British List can be found at BOU (2017)19. Category E has been compiled largely from county/regional bird reports and other published listings, and is not exhaustive. BOURC is unable to confirm the correct identification of species in Category E, or the circumstances in which they were recorded. Doubtful cases have been excluded, pending enquiries, along with species not recorded since 1 January 1950. The BOU is responsible for maintaining the British List. Part of this responsibility is to monitor the occurrence of non-native species which may qualify for addition to Category C of the British List. To undertake this we require published information from which to work and which we can quote as reference. The BOU encourages observers to report records of non-native species to the relevant local bird recorder. Local recorders and those producing local, county or regional publications are encouraged to publish these records, and to make them available to the Rare Breeding Birds Panel (RBBP) who publish annual reports1- 7,9,10,12 of non-native species breeding in Britain in British Birds. -

Glossy Ibis and the Identification Challenges of Hybrid Plegadis in Colorado
IN THE SCOPE Glossy Ibis and the identification challenges of hybrid Plegadis in Colorado Tony Leukering Glossy Ibis (Plegadis falcinellus) has, in the past 20 years or so, be- come a regular, though still rare, part of Colorado’s migrant avifauna. The CBRC lists 39 accepted records and though the first record was from Delta on 27 March 1986, virtually all other records are from the Eastern Plains and all but three (including the above) occurred in the period 11 April - 28 May. Though the rate of accrual of accepted records is at the threshold at which the species may be removed from the state review list (Anonymous 2002), the Colorado Bird Records Committee has not removed it for two reasons. The first is that field identification of Glossy and White-faced (P. chihi) ibises is difficult, particularly for birds not in alternate plumage, and continued confu- sion in that vein certainly seems to me to be a valid cause for reten- tion of the species on the review list. The second is that the advent of apparent hybridization between the two species—including apparent hybrid individuals in Oklahoma (Arterburn and Grzybowski 2003) and apparent mixed breeding pairs in Wyoming (Faulkner 2005)— complicates identification of some alternate-plumaged adults. In fact, hybrid ibis have been reported from Colorado during each of the past four springs, with some of the birds that exhibit mixed characteristics being nicely photographed. I was lucky to find and photograph such a bird near Beebe Draw, Weld, on 23 April 2005 and pictures of it (the top two pictures presented on the back cover) provide the basis for this column. -

Sacred Ibis (Threskiornis Aethiopicus)1
Archival copy: for current recommendations see http://edis.ifas.ufl.edu or your local extension office. WEC267 Florida's Introduced Birds: Sacred Ibis (Threskiornis aethiopicus)1 Steve Johnson and Monica McGarrity2 Many non-native birds have been introduced in prey. Sacred Ibises are larger than Florida's native Florida—perhaps as many as 200 species! Of these, ibises. Adults are approximately 30 inches long (75 at least 16 introduced species are considered cm) with a wingspan of 44–49 inches (112–124 established, according to various authorities, and cm) and weigh about 3 pounds (1.4 kg). Sacred Ibises some are now considered invasive and could have (Fig. 1) have mostly white bodies and wings; the serious impacts in Florida. This fact sheet introduces trailing edges of their wings (tips of the feathers) are the Sacred Ibis, and is one of a series of fact sheets gray-black. They have very distinctive long, black about Florida's established non-native birds and their feathers or plumes on their rumps (Fig. 1). During the impacts on our native ecosystems, economy, and the breeding season the feathers on the sides of their quality of life of Floridians. For more information on chests and on the outer wings (near the edge when Florida's introduced birds, how they got here, and the folded) may have a yellowish (or reddish) tinge, and problems they cause, read "Florida's Introduced their lower legs may be tinged with reddish-copper; Birds: An Overview" bare patches of scarlet-red skin may also be visible (http://edis.ifas.ufl.edu/UW297) and the other fact under their wings. -

First Record of Buff-Necked Ibis Theristicus Caudatus for Peru
Cotinga 33 First record of Buff-necked Ibis Theristicus caudatus for Peru Robert S. R. Williams, Maura Angelica Jurado Zevallos, Ernesto Fernandez Gamarra and Leoncio Flores Balarezo Received 4 March 2011; final revision accepted 26 April 2011 Cotinga 33 (2011): 92–93 Presentamos el primer registro de la Bandurria de Ala Blanca Theristicus caudatus para el Perú. Registramos tres individuos en las Pampas del Heath, Parque Nacional Bajuaja-Sonene, dpto. Puno en septiembre de 2009 y dos individuos en el mismo localidad en agosto de 2010. El registro fue documentado con fotos y grabaciones de sonidos. Buff-necked Ibis Theristicus caudatus is a polytypic the río Heath and Quebrada Juliaca. This is the species of lowland savannas and wetlands in same area visited by Zeppilli et al.5 in 2006 and that South America: T. c. caudatus occurs in Colombia most visited by other biologists, it being the most and Venezuela, and T. c. hyperorius south of the accessible savanna habitat. It forms part of the Amazon in Brazil, Bolivia, Paraguay, Uruguay and 1,091,416-ha Bahuaja-Sonene National Park, in Argentina4. In Bolivia, the species is widespread in extreme south-east Peru (dptos. Madre de Dios and the lowlands and foothills of the Andes to 2,400 m in Puno). The Pampas del Heath is the only edaphic dptos. Beni, Cochabamba, Santa Cruz, Chuquisaca, tropical savanna in Peru, with an area of 6,549 ha Tarija and La Paz, where this subspecies reaches (based on a Landsat image from 24 August 2010) its north-west limit at Pilón Lajas3.