An Oceanographic, Meteorological, and Biological 'Perfect Storm'yields
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Trout Fishing in Hillsdale July, 2012 Guy Winig, Member Hillsdale Conservation Advisory Council
Roe Jan Brown-released to fight another day. Trout Fishing in Hillsdale July, 2012 Guy Winig, member Hillsdale Conservation Advisory Council Trout are our canaries in a coalmine. They are an excellent indicator of stream health, requiring clean, cold, well-oxygenated water to thrive. Without very high quality surface waters, trout just cannot survive. Hillsdale is fortunate to have several miles of trout streams including the Roeliff Jansen Kill, Taconic Creek, Agawamuck Creek and Green River with numerous smaller and unnamed tributaries contributing to them. Our streams and their residents endure many perils including natural flood and drought cycles as well as man made impacts like pollution, invasive species, barriers and over –fishing. Erosion adds silt that smothers aquatic food sources and gravel spawning beds. Introduced Didymo algae (a.k.a. ‘Rock Snot’) now found in the Green River, also chokes the streambed. Loss of vegetation exposes water to heating and eliminates important breeding, feeding and cover areas for trout. Readers may recall the massive fish kill that resulted from the release of manure into the Roe Jan in the 90’s or tales of unscrupulous ‘sportsmen’ using small explosives and chlorine to kill all the fish in a segment of stream. Maybe the most harmful and insidious are non-point sources of pollution such as chemicals, fertilizers and heated runoff from parking lots, roads, roofs, crop fields and lawns. In 2009 and 2010, members of the community with help from the Columbia-Greene chapter of Trout Unlimited and the N.Y.S.D.E.C. Trees for Tributaries program planted several hundred small trees and shrubs on the banks and flood plain of the Roe Jan Kill through the Roe Jan Park in an effort to stabilize banks, introduce vegetative cover and provide shade to maintain cool stream temperatures. -

New and Emerging Technologies for Sustainable Fisheries: a Comprehensive Landscape Analysis
Photo by Pablo Sanchez Quiza New and Emerging Technologies for Sustainable Fisheries: A Comprehensive Landscape Analysis Environmental Defense Fund | Oceans Technology Solutions | April 2021 New and Emerging Technologies for Sustainable Fisheries: A Comprehensive Landscape Analysis Authors: Christopher Cusack, Omisha Manglani, Shems Jud, Katie Westfall and Rod Fujita Environmental Defense Fund Nicole Sarto and Poppy Brittingham Nicole Sarto Consulting Huff McGonigal Fathom Consulting To contact the authors please submit a message through: edf.org/oceans/smart-boats edf.org | 2 Contents List of Acronyms ...................................................................................................................................................... 5 1. Introduction .............................................................................................................................................................7 2. Transformative Technologies......................................................................................................................... 10 2.1 Sensors ........................................................................................................................................................... 10 2.2 Satellite remote sensing ...........................................................................................................................12 2.3 Data Collection Platforms ...................................................................................................................... -

Marine Fish Conservation Global Evidence for the Effects of Selected Interventions
Marine Fish Conservation Global evidence for the effects of selected interventions Natasha Taylor, Leo J. Clarke, Khatija Alliji, Chris Barrett, Rosslyn McIntyre, Rebecca0 K. Smith & William J. Sutherland CONSERVATION EVIDENCE SERIES SYNOPSES Marine Fish Conservation Global evidence for the effects of selected interventions Natasha Taylor, Leo J. Clarke, Khatija Alliji, Chris Barrett, Rosslyn McIntyre, Rebecca K. Smith and William J. Sutherland Conservation Evidence Series Synopses 1 Copyright © 2021 William J. Sutherland This work is licensed under a Creative Commons Attribution 4.0 International license (CC BY 4.0). This license allows you to share, copy, distribute and transmit the work; to adapt the work and to make commercial use of the work providing attribution is made to the authors (but not in any way that suggests that they endorse you or your use of the work). Attribution should include the following information: Taylor, N., Clarke, L.J., Alliji, K., Barrett, C., McIntyre, R., Smith, R.K., and Sutherland, W.J. (2021) Marine Fish Conservation: Global Evidence for the Effects of Selected Interventions. Synopses of Conservation Evidence Series. University of Cambridge, Cambridge, UK. Further details about CC BY licenses are available at https://creativecommons.org/licenses/by/4.0/ Cover image: Circling fish in the waters of the Halmahera Sea (Pacific Ocean) off the Raja Ampat Islands, Indonesia, by Leslie Burkhalter. Digital material and resources associated with this synopsis are available at https://www.conservationevidence.com/ -

Ecosystem Services Generated by Fish Populations
AR-211 Ecological Economics 29 (1999) 253 –268 ANALYSIS Ecosystem services generated by fish populations Cecilia M. Holmlund *, Monica Hammer Natural Resources Management, Department of Systems Ecology, Stockholm University, S-106 91, Stockholm, Sweden Abstract In this paper, we review the role of fish populations in generating ecosystem services based on documented ecological functions and human demands of fish. The ongoing overexploitation of global fish resources concerns our societies, not only in terms of decreasing fish populations important for consumption and recreational activities. Rather, a number of ecosystem services generated by fish populations are also at risk, with consequences for biodiversity, ecosystem functioning, and ultimately human welfare. Examples are provided from marine and freshwater ecosystems, in various parts of the world, and include all life-stages of fish. Ecosystem services are here defined as fundamental services for maintaining ecosystem functioning and resilience, or demand-derived services based on human values. To secure the generation of ecosystem services from fish populations, management approaches need to address the fact that fish are embedded in ecosystems and that substitutions for declining populations and habitat losses, such as fish stocking and nature reserves, rarely replace losses of all services. © 1999 Elsevier Science B.V. All rights reserved. Keywords: Ecosystem services; Fish populations; Fisheries management; Biodiversity 1. Introduction 15 000 are marine and nearly 10 000 are freshwa ter (Nelson, 1994). Global capture fisheries har Fish constitute one of the major protein sources vested 101 million tonnes of fish including 27 for humans around the world. There are to date million tonnes of bycatch in 1995, and 11 million some 25 000 different known fish species of which tonnes were produced in aquaculture the same year (FAO, 1997). -

Localized Plankton Blooms and Jubilees on the Gulf Coast Gordon Gunter Gulf Coast Research Laboratory
Gulf Research Reports Volume 6 | Issue 3 January 1979 Localized Plankton Blooms and Jubilees on the Gulf Coast Gordon Gunter Gulf Coast Research Laboratory Charles H. Lyles Gulf Coast Research Laboratory DOI: 10.18785/grr.0603.12 Follow this and additional works at: http://aquila.usm.edu/gcr Part of the Marine Biology Commons Recommended Citation Gunter, G. and C. H. Lyles. 1979. Localized Plankton Blooms and Jubilees on the Gulf Coast. Gulf Research Reports 6 (3): 297-299. Retrieved from http://aquila.usm.edu/gcr/vol6/iss3/12 This Short Communication is brought to you for free and open access by The Aquila Digital Community. It has been accepted for inclusion in Gulf and Caribbean Research by an authorized editor of The Aquila Digital Community. For more information, please contact [email protected]. Gulf Research Reports, Vol. 6,NO. 3,291-299, 1919. LOCALIZED PLANKTON BLOOMS AND JUBILEES ON THE GULF COAST GORDON GUNTER AND CHARLES H. LYLES Gulf Coast Research Laboratory, Ocean Springs, Mississippi 39564 and Gulf States Marine Fisheries Commission, Ocean Springs, Mississippi 39564 ABSTRACT The writers describe various small types of plankton blooms such as those occurring in boat slips, the head of a large bayou and a strip type bloom of Chaeroceras on the Gulf beach. Oyster kills from “poison water” draining off of marshes are said to be caused by plankton bloom. Small “jubilees” are said to be caused by localized blooms and one of these is described as it occurred. In November 1938, Dr. Margaretha Brongersma-Sanders of billions of Chaetoceras sp., a common open sea diatom. -

Fish Passage Engineering Design Criteria 2019
FISH PASSAGE ENGINEERING DESIGN CRITERIA 2019 37.2’ U.S. Fish and Wildlife Service Northeast Region June 2019 Fish and Aquatic Conservation, Fish Passage Engineering Ecological Services, Conservation Planning Assistance United States Fish and Wildlife Service Region 5 FISH PASSAGE ENGINEERING DESIGN CRITERIA June 2019 This manual replaces all previous editions of the Fish Passage Engineering Design Criteria issued by the U.S. Fish and Wildlife Service Region 5 Suggested citation: USFWS (U.S. Fish and Wildlife Service). 2019. Fish Passage Engineering Design Criteria. USFWS, Northeast Region R5, Hadley, Massachusetts. USFWS R5 Fish Passage Engineering Design Criteria June 2019 USFWS R5 Fish Passage Engineering Design Criteria June 2019 Contents List of Figures ................................................................................................................................ ix List of Tables .................................................................................................................................. x List of Equations ............................................................................................................................ xi List of Appendices ........................................................................................................................ xii 1 Scope of this Document ....................................................................................................... 1-1 1.1 Role of the USFWS Region 5 Fish Passage Engineering ............................................ -

Symposium Review: Using Electronic Tags to Estimate Vital Rates in Fishes Brendan J
COLUMN 2017 ANNUAL MEETING Symposium Review: Using Electronic Tags to Estimate Vital Rates in Fishes Brendan J. Runde | Center for Marine Sciences and Technology, Department of Applied Ecology, North Carolina State University, 303 College Cir., Morehead City, NC 28557. E-mail: [email protected] Julianne E. Harris | U.S. Fish and Wildlife Service Office, Columbia River Fish and Wildlife Conservation, Vancouver, WA Jeffrey A. Buckel | Center for Marine Sciences and Technology, Department of AppliedEcology, North Carolina State University, Morehead City, NC Accurate and precise estimates of vital rates, such as nat- discussion that concluded the session. Symposium presenta- ural and fishing mortality, can be challenging to obtain, but tions illustrated the breadth and depth of use of electronic are essential for stock assessment, fisheries management, and tags to estimate vital rates and the discussion stressed the conservation. Joseph Hightower pioneered a novel approach need for careful study design. As tag technology and ana- for estimating mortality rates—assigning fates of electroni- lytical approaches continue to improve, we suggest that elec- cally tagged fish using telemetry. Herein, we describe a day- tronic tagging studies will become even more valuable to help long symposium held at the 147th American Fisheries Society scientists and managers better understand fish biology and Meeting in 2017 dedicated to highlighting and discussing manage fisheries. current uses of and recent advancements in electronic tag- Accurate and precise estimates of vital rates such as fishing ging to estimate fish vital rates. In this review, we highlight mortality (F) and natural mortality (M) are necessary for stock the 18 presentations made in the symposium as well as the assessment, fisheries management, and conservation. -

Little Fish, Big Impact: Managing a Crucial Link in Ocean Food Webs
little fish BIG IMPACT Managing a crucial link in ocean food webs A report from the Lenfest Forage Fish Task Force The Lenfest Ocean Program invests in scientific research on the environmental, economic, and social impacts of fishing, fisheries management, and aquaculture. Supported research projects result in peer-reviewed publications in leading scientific journals. The Program works with the scientists to ensure that research results are delivered effectively to decision makers and the public, who can take action based on the findings. The program was established in 2004 by the Lenfest Foundation and is managed by the Pew Charitable Trusts (www.lenfestocean.org, Twitter handle: @LenfestOcean). The Institute for Ocean Conservation Science (IOCS) is part of the Stony Brook University School of Marine and Atmospheric Sciences. It is dedicated to advancing ocean conservation through science. IOCS conducts world-class scientific research that increases knowledge about critical threats to oceans and their inhabitants, provides the foundation for smarter ocean policy, and establishes new frameworks for improved ocean conservation. Suggested citation: Pikitch, E., Boersma, P.D., Boyd, I.L., Conover, D.O., Cury, P., Essington, T., Heppell, S.S., Houde, E.D., Mangel, M., Pauly, D., Plagányi, É., Sainsbury, K., and Steneck, R.S. 2012. Little Fish, Big Impact: Managing a Crucial Link in Ocean Food Webs. Lenfest Ocean Program. Washington, DC. 108 pp. Cover photo illustration: shoal of forage fish (center), surrounded by (clockwise from top), humpback whale, Cape gannet, Steller sea lions, Atlantic puffins, sardines and black-legged kittiwake. Credits Cover (center) and title page: © Jason Pickering/SeaPics.com Banner, pages ii–1: © Brandon Cole Design: Janin/Cliff Design Inc. -

FISHING NEWSLETTER 2020/2021 Table of Contents FWP Administrative Regions and Hatchery Locations
FISHING NEWSLETTER 2020/2021 Table of Contents FWP Administrative Regions and Hatchery Locations .........................................................................................3 Region 1 Reports: Northwest Montana ..........................................................................................................5 Region 2 Reports: West Central Montana .....................................................................................................17 Region 3 Reports: Southwest Montana ........................................................................................................34 Region 4 Reports: North Central Montana ...................................................................................................44 Region 5 Reports: South Central Montana ...................................................................................................65 Region 6 Reports: Northeast Montana ........................................................................................................73 Region 7 Reports: Southeast Montana .........................................................................................................86 Montana Fish Hatchery Reports: .......................................................................................................................92 Murray Springs Trout Hatchery ...................................................................................................................92 Washoe Park Trout Hatchery .......................................................................................................................93 -

Catch and Release Fishing; Effects on Bass Populations
Pennsylvania Fish and Boat Commission Harrisburg PA www.fish.state.pa.us Catch & Release Fishing; Effects on Bass Populations By PFBC Staff Catch-and-release fishing is often seen by the public as an effective way of keeping good-sized largemouth bass and smallmouth bass in a population so that these fish can be caught again. But does it work? That is, how well do bass survive catch and release, and are there things anglers can do to improve the odds? A lot of research has been conducted on this topic. Here we will discuss the facts of catch-and-release mortality and how anglers can help improve bass survival. Catch-and-release fishing has become the norm, instead of the exception. Anglers proudly proclaim they release their catch with bumper stickers, hats and t-shirts. But just because the bass swims away, its survival isn’t guaranteed. Many anglers wonder about bass survival after fishing tourna- ments, and if catch and release works to keep good-sized bass in their local lakes and rivers. Much research on this topic has been done in Pennsylvania and elsewhere. There are many issues that affect if a bass will survive being caught or not. By using the best practices possible, anglers can help bass survive. Mortality Mortality can come about two ways in fish; naturally and through fishing. Natural causes include disease or being eaten by a predator. Fishing mortality can come from either the fish being harvested by an angler or from injuries or stress from catch and release. The mortality rate of a waterway is a measure of how many fish die or are removed over a year. -
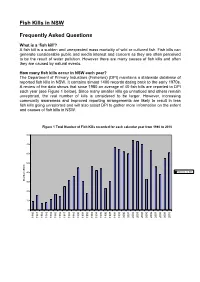
Fish Kills in NSW Frequently Asked Questions
Fish Kills in NSW Frequently Asked Questions What is a ‘fish kill’? A fish kill is a sudden and unexpected mass mortality of wild or cultured fish. Fish kills can generate considerable public and media interest and concern as they are often perceived to be the result of water pollution. However there are many causes of fish kills and often they are caused by natural events. How many fish kills occur in NSW each year? The Department of Primary Industries (Fisheries) (DPI) maintains a statewide database of reported fish kills in NSW. It contains almost 1400 records dating back to the early 1970s. A review of the data shows that since 1980 an average of 40 fish kills are reported to DPI each year (see Figure 1 below). Since many smaller kills go unnoticed and others remain unreported, the real number of kills is considered to be larger. However, increasing community awareness and improved reporting arrangements are likely to result in less fish kills going unreported and will also assist DPI to gather more information on the extent and causes of fish kills in NSW. Figure 1 Total Number of Fish Kills recorded for each calendar year from 1980 to 2010 80 70 60 50 40 Total no. of kills Number of kills 30 20 10 0 1980 1981 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 What species of fish are affected? Over 100 species of fish, including finfish, molluscs and crustaceans, have been reported in fish kills in NSW since records began. -

Proposed Incidental Harassment Authorizations for Seismic Surveys in the Atlantic Ocean
AMERICAN LITTORAL SOCIETY – ANIMAL LEGAL DEFENSE FUND – ANIMAL WELFARE INSTITUTE – CAPE FEAR RIVER WATCH – CENTER FOR A SUSTAINABLE COAST – CENTER FOR BIOLOGICAL DIVERSITY CET LAW – CLEAN OCEAN ACTION – COASTAL CONSERVATION LEAGUE – CONSERVATION LAW FOUNDATION – COOK INLETKEEPER – DEFENDERS OF WILDLIFE – THE DOLPHIN PROJECT – EARTH LAW CENTER – EARTHJUSTICE – ENVIRONMENT GEORGIA – THE HUMANE SOCIETY OF THE U.S. – INITIATIVE TO PROTECT JEKYLL ISLAND – INTERNATIONAL FUND FOR ANIMAL WELFARE – MATANZAS RIVERKEEPER – MIAMI WATERKEEPER – NATURAL RESOURCES DEFENSE COUNCIL – NORTH CAROLINA COASTAL FEDERATION – NORTH CAROLINA WILDLIFE FEDERATION – OCEAN CONSERVATION RESEARCH – ONE HUNDRED MILES – ST. JOHNS RIVERKEEPER – SATILLA RIVERKEEPER – SIERRA CLUB – SIERRA CLUB ATLANTIC CHAPTER – SIERRA CLUB CHAPTERS OF FLORIDA, GEORGIA, MAINE, MARYLAND, MASSACHUSETTS, NEW JERSEY, NORTH CAROLINA, SOUTH CAROLINA, AND VIRGINIA – SOUND RIVERS – SOUTH CAROLINA WILDLIFE FEDERATION – SOUTHERN ENVIRONMENTAL LAW CENTER – STOP OFFSHORE DRILLING IN THE ATLANTIC (SODA) – SURFRIDER FOUNDATION – WHALE AND DOLPHIN CONSERVATION – WORLD WILDLIFE FUND PUBLIC HEARINGS REQUESTED By Electronic Mail July 21, 2017 Ms. Jolie Harrison Chief, Permits and Conservation Division Office of Protected Resources National Marine Fisheries Service 1315 East-West Highway Silver Spring, MD 20910 [email protected] Re: Proposed incidental harassment authorizations for seismic surveys in the Atlantic Ocean Dear Ms. Harrison: On behalf of the Natural Resources Defense Council (“NRDC”), Center