New Insights in Citron (Citrus Medica L.) Genomics and Fruit Development Eliezer E
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Kettleman City, Kings County Please Read Immediately
CALIFORNIA DEPARTMENT OF FOOD AND AGRICULTURE OFFICIAL NOTICE FOR CITY OF KETTLEMAN CITY, KINGS COUNTY PLEASE READ IMMEDIATELY THE NOTICE OF TREATMENT FOR THE ASIAN CITRUS PSYLLID On September 18, 2020 the California Department of Food and Agriculture (CDFA) confirmed the presence of Asian citrus psyllid (ACP), Diaphorina citri Kuwayama, a harmful exotic pest in the city of Kettleman City, Kings County. This detection indicate that a breeding population exists in the area. The devastating citrus disease Huanglongbing (HLB) is spread by the feeding action of ACP. The ACP infestation is sufficiently isolated and localized to be amenable to the CDFA’s ACP treatment work plan. A Program Environmental Impact Report (PEIR) has been certified which analyzes the ACP treatment program in accordance with Public Resources Code, Sections 21000 et seq. The PEIR is available at http://www.cdfa.ca.gov/plant/peir/. The treatment activities described below are consistent with the PEIR. In accordance with integrated pest management principles, CDFA has evaluated possible treatment methods and determined that there are no physical, cultural, or biological control methods available to eliminate the ACP from this area. Notice of Treatment is valid until September 18, 2021, which is the amount of time necessary to determine that the treatment was successful. The treatment plan for the ACP infestation will be implemented within a 50-meter radius of each detection site, as follows: • Tempo® SC Ultra (cyfluthrin), a contact insecticide for controlling the adults and nymphs of ACP, will be applied from the ground using hydraulic spray equipment to the foliage of host plants; and • Merit® 2F or CoreTect™ (imidacloprid), a systemic insecticide for controlling the immature life stages of ACP, will be applied to the soil underneath host plants. -

Texas Citrus Tree Valuesjuan R
EHT-011 7/13 Texas Citrus Tree ValuesJuan R. Anciso and Luis A. Ribera* Photo courtesy of Rod Santa Ana ommercial citrus growers and homeowners To figure the total value per tree for years 2 to often need to determine the value of their 6, add the value of a tree the previous year plus mature citrus trees. The objective is to deter- the potential gross revenue the new tree is bring- mine the value of a mature grapefruit tree (Rio ing. RedC variety) and a mature orange tree (Valencia Finally, calculate the total value of mature variety) to assess the value of a tree or an orchard. trees (7 years and older) by adding the cumula- Grapefruit and orange trees usually reach full tive potential net revenue of a mature tree over 7 maturity by year 7 with an estimated yield of 23 years, plus the planting cost minus the cumulative and 18 tons per acre, respectively (Table 1.) How- potential net revenue of a new tree, years 1 to 6. ever, some production is expected from year 2 Overall, citrus tree values vary from year 1 with yearly increments until they reach maturity. through 7. At year 7, citrus trees are fully mature The price per ton used in this study is the due to their commercial production potential. Of 5-year average (2008–2012) producers received course, the longevity or lifespan of a citrus tree for either grapefruit or Valencia oranges. The cost depends on its care and whether it endures any of production is separated into planting costs or type of cold injury and rehabilitation, but the esti- establishment costs and annual production cost. -

A History of Fruits on the Southeast Asian Mainland
OFFPRINT A history of fruits on the Southeast Asian mainland Roger Blench Kay Williamson Educational Foundation Cambridge, UK E-mail: [email protected] http://www.rogerblench.info/RBOP.htm Occasional Paper 4 Linguistics, Archaeology and the Human Past Edited by Toshiki OSADA and Akinori UESUGI Indus Project Research Institute for Humanity and Nature, Kyoto, Japan 2008 ISBN 978-4-902325-33-1 A history of Fruits on the Southeast Asian mainland A history of fruits on the Southeast Asian mainland Roger Blench Kay Williamson Educational Foundation Cambridge, UK E-mail: [email protected] http://www.rogerblench.info/RBOP.htm ABSTRACT The paper presents an overview of the history of the principal tree fruits grown on the Southeast Asian mainland, making use of data from biogeography, archaeobotany, iconography and linguistics. Many assertions in the literature about the origins of particular species are found to be without empirical basis. In the absence of other data, comparative linguistics is an important source for tracing the spread of some fruits. Contrary to the Pacific, it seems that many of the fruits we now consider characteristic of the region may well have spread in recent times. INTRODUCTION empirical base for Pacific languages is not matched for mainland phyla such as Austroasiatic, Daic, Sino- This study 1) is intended to complement a previous Tibetan or Hmong-Mien, so accounts based purely paper on the history of tree-fruits in island Southeast on Austronesian tend to give a one-sided picture. Asia and the Pacific (Blench 2005). Arboriculture Although occasional detailed accounts of individual is very neglected in comparison to other types of languages exist (e.g. -

Bright Citron™ Soak
Product Profile BRIGHT CITRON™ SOAK WHAT IT IS An aromatic and refreshing sea salt soak for cleansing skin. WHAT IT DOES Softens and cleanses skin. WHY YOU NEED IT • Provides the first step to cleanse impurities and soften skin for a luxurious manicure or pedicure experience. • Gently cleanses without drying skin. FRAGRANCE FEATURED Pink Grapefruit and Warm Amber. FRAGRANCE PROFILE • Bright citrus top notes. • Soft floral mid notes. CREATIVE SUGGESTIONS • Light woody base notes. • Create a fragrant ceremonial experience by placing the Soak into warm water directly prior to ACTIVE BOTANICALS immersing hands or feet. Kaffir Lime (Citrus Hystrix Leaf Extract): • Add lime or grapefruit slices to enhance the Is known to purify skin and promote radiance. experience. • Use as a foot soak prior to all salon services. Honey: • Excellent retail product for home use in bath tubs. Is known to hydrate skin and help retain moisture. INGREDIENT LISTING FEATURED INGREDIENTS & BENEFITS Maris Sal ((Sea Salt) Sel Marin), Sodium Sesquicarbonate, Sea Salt: Argania Spinosa Kernel Oil, Honey, Citrus Hystrix Leaf Natural salt used for its therapeutic properties. Extract, Aqua ((Water) Eau), Glycerin, Propylene Glycol, Salts increase the flow of water in and out of Hydrated Silica, Parfum (Fragrance), Limonene, Linalool, cells, in essence flushing and cleansing the cells. Hexyl Cinnamal, Citral. This facilitates the absorption of other ingredients into the cells. AVAILABLE SIZES • Retail/Professional: 410 g (14.4 oz) Argan Oil (Argania Spinosa Kernel Oil): • Professional: 3.3 kg (118.8 oz) Is known to nourish dry skin. DIRECTIONS FOR USE • For feet, add a teaspoon to footbath and agitate water to dissolve. -

Tropical Horticulture: Lecture 32 1
Tropical Horticulture: Lecture 32 Lecture 32 Citrus Citrus: Citrus spp., Rutaceae Citrus are subtropical, evergreen plants originating in southeast Asia and the Malay archipelago but the precise origins are obscure. There are about 1600 species in the subfamily Aurantioideae. The tribe Citreae has 13 genera, most of which are graft and cross compatible with the genus Citrus. There are some tropical species (pomelo). All Citrus combined are the most important fruit crop next to grape. 1 Tropical Horticulture: Lecture 32 The common features are a superior ovary on a raised disc, transparent (pellucid) dots on leaves, and the presence of aromatic oils in leaves and fruits. Citrus has increased in importance in the United States with the development of frozen concentrate which is much superior to canned citrus juice. Per-capita consumption in the US is extremely high. Citrus mitis (calamondin), a miniature orange, is widely grown as an ornamental house pot plant. History Citrus is first mentioned in Chinese literature in 2200 BCE. First citrus in Europe seems to have been the citron, a fruit which has religious significance in Jewish festivals. Mentioned in 310 BCE by Theophrastus. Lemons and limes and sour orange may have been mutations of the citron. The Romans grew sour orange and lemons in 50–100 CE; the first mention of sweet orange in Europe was made in 1400. Columbus brought citrus on his second voyage in 1493 and the first plantation started in Haiti. In 1565 the first citrus was brought to the US in Saint Augustine. 2 Tropical Horticulture: Lecture 32 Taxonomy Citrus classification based on morphology of mature fruit (e.g. -

Cooperatives in the U.S.-Citrus Industry
Agriculture Cooperatives in the Rural Business and Cooperative Development U.S.-Citrus Industry Service RBCDS Research Report 137 Abstract Cooperatives in the U.S. Citrus Industry James A. Jacobs Agricultural Economist U.S. Department of Agriculture Rural Business and Cooperative Development Service Citrus is one of the leading fruit crops produced in the United States. Cooperatives play an important role in the handling and marketing of both fresh and processed citrus products. This report examines the development and posi- tion of cooperatives in the citrus industry, their functions and operating prac- tices, and the impact of changes in production practices and industry structure on cooperatives. Cooperatives range from small, local fresh packinghouse associations to large cooperative federations with comprehensive marketing and sales pro- grams in both fresh and processed markets. Cooperatives are among the lead- ing marketers in all producing areas, and are the dominant marketing organiza- tion in California and Arizona. Citrus cooperatives use the pooling method to market and allocate returns. This cooperative practice of averaging price and sharing risk is commonly used by some private citrus firms as well, reflecting the inherent volatility of citrus production. Keywords: Cooperative, grove, grower-member, fresh citrus, processed citrus, frozen-concentrated orange juice, packinghouse, processor, marketing federa- tion, sales agency, marketing agreement, pooling, grove care, freezes, box, eliminations. RBCDS Research Report 137 December 1994 Preface This report describes the position and functions of cooperatives in the U.S. citrus industry. It is the first known detailed examination of its kind on citrus cooperative activities and operating practices. The report is intended as a reference for cooperative managers and mem- bers, professional advisors, and anyone involved in professional activities or research in the citrus industry. -
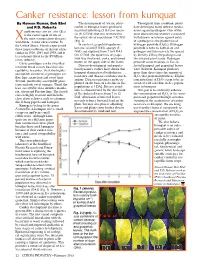
Canker Resistance: Lesson from Kumquat by Naveen Kumar, Bob Ebel the Development of Asiatic Citrus Throughout Their Evolution, Plants and P.D
Canker resistance: lesson from kumquat By Naveen Kumar, Bob Ebel The development of Asiatic citrus Throughout their evolution, plants and P.D. Roberts canker in kumquat leaves produced have developed many defense mecha- anthomonas citri pv. citri (Xcc) localized yellowing (5 DAI) or necro- nisms against pathogens. One of the is the causal agent of one of sis (9-12 DAI) that was restricted to most characteristic features associated the most serious citrus diseases the actual site of inoculation 7-12 DAI with disease resistance against entry X (Fig. 2). of a pathogen is the production of worldwide, Asiatic citrus canker. In the United States, Florida experienced In contrast, grapefruit epidermis hydrogen peroxide (H2O2). Hydrogen three major outbreaks of Asiatic citrus became raised (5 DAI), spongy (5 peroxide is toxic to both plant and canker in 1910, 1984 and 1995, and it DAI) and ruptured from 7 to 8 DAI. pathogen and thus restricts the spread is a constant threat to the $9 billion On 12 DAI, the epidermis of grape- by directly killing the pathogen and citrus industry. fruit was thickened, corky, and turned the infected plant tissue. Hydrogen Citrus genotypes can be classified brown on the upper side of the leaves. peroxide concentrations in Xcc-in- into four broad classes based on sus- Disease development and popula- fected kumquat and grapefruit leaves ceptibility to canker. First, the highly- tion dynamics studies have shown that were different. Kumquat produces susceptible commercial genotypes are kumquat demonstrated both disease more than three times the amount of Key lime, grapefruit and sweet lime. -

Citrus from Seed?
Which citrus fruits will come true to type Orogrande, Tomatera, Fina, Nour, Hernandina, Clementard.) from seed? Ellendale Tom McClendon writes in Hardy Citrus Encore for the South East: Fortune Fremont (50% monoembryonic) “Most common citrus such as oranges, Temple grapefruit, lemons and most mandarins Ugli Umatilla are polyembryonic and will come true to Wilking type. Because most citrus have this trait, Highly polyembryonic citrus types : will mostly hybridization can be very difficult to produce nucellar polyembryonic seeds that will grow true to type. achieve…. This unique characteristic Citrus × aurantiifolia Mexican lime (Key lime, West allows amateurs to grow citrus from seed, Indian lime) something you can’t do with, say, Citrus × insitorum (×Citroncirus webberii) Citranges, such as Rusk, Troyer etc. apples.” [12*] Citrus × jambhiri ‘Rough lemon’, ‘Rangpur’ lime, ‘Otaheite’ lime Monoembryonic (don’t come true) Citrus × limettioides Palestine lime (Indian sweet lime) Citrus × microcarpa ‘Calamondin’ Meyer Lemon Citrus × paradisi Grapefruit (Marsh, Star Ruby, Nagami Kumquat Redblush, Chironja, Smooth Flat Seville) Marumi Kumquat Citrus × sinensis Sweet oranges (Blonde, navel and Pummelos blood oranges) Temple Tangor Citrus amblycarpa 'Nasnaran' mandarin Clementine Mandarin Citrus depressa ‘Shekwasha’ mandarin Citrus karna ‘Karna’, ‘Khatta’ Poncirus Trifoliata Citrus kinokuni ‘Kishu mandarin’ Citrus lycopersicaeformis ‘Kokni’ or ‘Monkey mandarin’ Polyembryonic (come true) Citrus macrophylla ‘Alemow’ Most Oranges Citrus reshni ‘Cleopatra’ mandarin Changshou Kumquat Citrus sunki (Citrus reticulata var. austera) Sour mandarin Meiwa Kumquat (mostly polyembryonic) Citrus trifoliata (Poncirus trifoliata) Trifoliate orange Most Satsumas and Tangerines The following mandarin varieties are polyembryonic: Most Lemons Dancy Most Limes Emperor Grapefruits Empress Tangelos Fairchild Kinnow Highly monoembryonic citrus types: Mediterranean (Avana, Tardivo di Ciaculli) Will produce zygotic monoembryonic seeds that will not Naartje come true to type. -

Survey of Phenolic Compounds Produced in Citrus
USDA ??:-Z7 S rveyof Phenolic United States Department of Agriculture C mpounds Produced IliIIiI Agricultural Research In Citrus Service Technical Bulletin Number 1856 December 1998 United States Department of Agriculture Survey of Phenolic Compounds Agricultural Produced in Citrus Research Service Mark Berhow, Brent Tisserat, Katherine Kanes, and Carl Vandercook Technical Bulletin Number 1856 December 1998 This research project was conducted at USDA, Agricultural Research Service, Fruit and Vegetable Chem istry laboratory, Pasadena, California, where Berhow was a research chemist, TIsserat was a research geneticist, Kanes was a research associate, and Vandercook, now retired, was a research chemist. Berhow and Tisserat now work at the USDA-ARS National Center for AgriCUltural Utilization Research, Peoria, Illinois, where Berhow is a research chemist and Tisserat is a research geneticist. Abstract Berhow, M., B. Tisserat, K. Kanes, and C. Vandercook. 1998. Survey of Mention of trade names or companies in this publication is solely for the Phenolic Compounds Produced in Citrus. U.S. Department ofAgriculture, purpose of providing specific information and does not imply recommenda Agricultural Research Service, Technical Bulletin No. 1856, 158 pp. tion or endorsement by the U. S. Department ofAgriculture over others not mentioned. A survey of phenolic compounds, especially flavanones and flavone and flavonol compounds, using high pressure liquid chromatography was While supplies last, single copies of this publication may be obtained at no performed in Rutaceae, subfamily Aurantioideae, representing 5 genera, cost from- 35 species, and 114 cultivars. The average number of peaks, or phenolic USDA, ARS, National Center for Agricultural Utilization Research compounds, occurring in citrus leaf, flavedo, albedo, and juice vesicles 1815 North University Street were 21, 17, 15, and 9.3, respectively. -

Citrus Industry Biosecurity Plan 2015
Industry Biosecurity Plan for the Citrus Industry Version 3.0 July 2015 PLANT HEALTH AUSTRALIA | Citrus Industry Biosecurity Plan 2015 Location: Level 1 1 Phipps Close DEAKIN ACT 2600 Phone: +61 2 6215 7700 Fax: +61 2 6260 4321 E-mail: [email protected] Visit our web site: www.planthealthaustralia.com.au An electronic copy of this plan is available through the email address listed above. © Plant Health Australia Limited 2004 Copyright in this publication is owned by Plant Health Australia Limited, except when content has been provided by other contributors, in which case copyright may be owned by another person. With the exception of any material protected by a trade mark, this publication is licensed under a Creative Commons Attribution-No Derivs 3.0 Australia licence. Any use of this publication, other than as authorised under this licence or copyright law, is prohibited. http://creativecommons.org/licenses/by-nd/3.0/ - This details the relevant licence conditions, including the full legal code. This licence allows for redistribution, commercial and non-commercial, as long as it is passed along unchanged and in whole, with credit to Plant Health Australia (as below). In referencing this document, the preferred citation is: Plant Health Australia Ltd (2004) Industry Biosecurity Plan for the Citrus Industry (Version 3.0 – July 2015). Plant Health Australia, Canberra, ACT. Disclaimer: The material contained in this publication is produced for general information only. It is not intended as professional advice on any particular matter. No person should act or fail to act on the basis of any material contained in this publication without first obtaining specific and independent professional advice. -

Citrus Fruits Information Compiled by Sunkist Growers
Commodity Fact Sheet Citrus Fruits Information compiled by Sunkist Growers How Produced – Citrus trees are propagated asexually mandarin orange and a lemon, are less acidic than traditional through a procedure known as grafting which fuses two lemons. different varieties of plants. In the case of citrus trees, one variety, the rootstock, is selected for Desert grapefruit are harvested October its hardiness and the other variety, the through March while summer grapefruit scion, is selected for its high-quality are available May through September. fruits. The rootstock, grown from a Specialty citrus include Melo Golds seed, is typically a two- to three-year- and Oro Blancos, grapefruit varieties old seedling while the scion is a bud that are popular with those preferring a from a mature tree. Through grafting, sweeter taste. Pummelos, or “Chinese” the scion fuses to the rootstock and grapefruit, considered a delicacy among becomes a new tree. In approximately many Asian cultures, are the largest of five years, the tree produces the same all citrus fruits. variety of fruit that was budded onto Almost a dozen different mandarin the rootstock. The successfully grafted and tangerine varieties, such as trees are sold to citrus growers through Clementines, Gold Nuggets, and wholesale nurseries and are certified Pixies, are available November through disease-free. There are approximately CITRUS VARIETIES May. Most are easy to peel and have a 270,000 bearing acres of citrus trees in lively flavor. California. Commodity Value – While Florida is the number one History – Oranges and lemons can be traced back to the producer of citrus fruits, the majority of their crop is made ancient Middle East. -
Holdings of the University of California Citrus Variety Collection 41
Holdings of the University of California Citrus Variety Collection Category Other identifiers CRC VI PI numbera Accession name or descriptionb numberc numberd Sourcee Datef 1. Citron and hybrid 0138-A Indian citron (ops) 539413 India 1912 0138-B Indian citron (ops) 539414 India 1912 0294 Ponderosa “lemon” (probable Citron ´ lemon hybrid) 409 539491 Fawcett’s #127, Florida collection 1914 0648 Orange-citron-hybrid 539238 Mr. Flippen, between Fullerton and Placentia CA 1915 0661 Indian sour citron (ops) (Zamburi) 31981 USDA, Chico Garden 1915 1795 Corsican citron 539415 W.T. Swingle, USDA 1924 2456 Citron or citron hybrid 539416 From CPB 1930 (Came in as Djerok which is Dutch word for “citrus” 2847 Yemen citron 105957 Bureau of Plant Introduction 3055 Bengal citron (ops) (citron hybrid?) 539417 Ed Pollock, NSW, Australia 1954 3174 Unnamed citron 230626 H. Chapot, Rabat, Morocco 1955 3190 Dabbe (ops) 539418 H. Chapot, Rabat, Morocco 1959 3241 Citrus megaloxycarpa (ops) (Bor-tenga) (hybrid) 539446 Fruit Research Station, Burnihat Assam, India 1957 3487 Kulu “lemon” (ops) 539207 A.G. Norman, Botanical Garden, Ann Arbor MI 1963 3518 Citron of Commerce (ops) 539419 John Carpenter, USDCS, Indio CA 1966 3519 Citron of Commerce (ops) 539420 John Carpenter, USDCS, Indio CA 1966 3520 Corsican citron (ops) 539421 John Carpenter, USDCS, Indio CA 1966 3521 Corsican citron (ops) 539422 John Carpenter, USDCS, Indio CA 1966 3522 Diamante citron (ops) 539423 John Carpenter, USDCS, Indio CA 1966 3523 Diamante citron (ops) 539424 John Carpenter, USDCS, Indio