All-D Amino Acid-Containing Channel-Forming Antibiotic Peptides
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
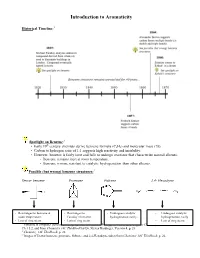
Introduction to Aromaticity
Introduction to Aromaticity Historical Timeline:1 Spotlight on Benzene:2 th • Early 19 century chemists derive benzene formula (C6H6) and molecular mass (78). • Carbon to hydrogen ratio of 1:1 suggests high reactivity and instability. • However, benzene is fairly inert and fails to undergo reactions that characterize normal alkenes. - Benzene remains inert at room temperature. - Benzene is more resistant to catalytic hydrogenation than other alkenes. Possible (but wrong) benzene structures:3 Dewar benzene Prismane Fulvene 2,4- Hexadiyne - Rearranges to benzene at - Rearranges to - Undergoes catalytic - Undergoes catalytic room temperature. Faraday’s benzene. hydrogenation easily. hydrogenation easily - Lots of ring strain. - Lots of ring strain. - Lots of ring strain. 1 Timeline is computer-generated, compiled with information from pg. 594 of Bruice, Organic Chemistry, 4th Edition, Ch. 15.2, and from Chemistry 14C Thinkbook by Dr. Steven Hardinger, Version 4, p. 26 2 Chemistry 14C Thinkbook, p. 26 3 Images of Dewar benzene, prismane, fulvene, and 2,4-Hexadiyne taken from Chemistry 14C Thinkbook, p. 26. Kekulé’s solution: - “snake bites its own tail” (4) Problems with Kekulé’s solution: • If Kekulé’s structure were to have two chloride substituents replacing two hydrogen atoms, there should be a pair of 1,2-dichlorobenzene isomers: one isomer with single bonds separating the Cl atoms, and another with double bonds separating the Cl atoms. • These isomers were never isolated or detected. • Rapid equilibrium proposed, where isomers interconvert so quickly that they cannot be isolated or detected. • Regardless, Kekulé’s structure has C=C’s and normal alkene reactions are still expected. - But the unusual stability of benzene still unexplained. -

Isomer Distributions of Molecular Weight 247 and 273 Nitro-Pahs in Ambient Samples, NIST Diesel SRM, and from Radical-Initiated Chamber Reactions
Atmospheric Environment 55 (2012) 431e439 Contents lists available at SciVerse ScienceDirect Atmospheric Environment journal homepage: www.elsevier.com/locate/atmosenv Isomer distributions of molecular weight 247 and 273 nitro-PAHs in ambient samples, NIST diesel SRM, and from radical-initiated chamber reactions Kathryn Zimmermann a,1, Roger Atkinson a,1,2,3, Janet Arey a,1,2,*, Yuki Kojima b,4, Koji Inazu b,5 a Air Pollution Research Center, University of California, Riverside, CA 92521, USA b Tokyo Institute of Technology, 4259 Nagatsuta, Midori-ku, Yokohama 226-8502, Japan article info abstract Article history: Molecular weight (mw) 247 nitrofluoranthenes and nitropyrenes and mw 273 nitrotriphenylenes (NTPs), Received 27 December 2011 nitrobenz[a]anthracenes, and nitrochrysenes were quantified in ambient particles collected in Riverside, Received in revised form CA, Tokyo, Japan, and Mexico City, Mexico. 2-Nitrofluoranthene (2-NFL) was the most abundant nitro- 28 February 2012 polycyclic aromatic hydrocarbon (nitro-PAH) in Riverside and Mexico City, and the mw 273 nitro-PAHs Accepted 5 March 2012 were observed in lower concentrations. However, in Tokyo concentrations of 1- þ 2-NTP were more similar to that of 2-NFL. NIST SRM 1975 diesel extract standard reference material was also analyzed to Keywords: examine nitro-PAH isomer distributions, and 12-nitrobenz[a]anthracene was identified for the first time. Nitro-PAH fl Atmospheric reactions The atmospheric formation pathways of nitro-PAHs were studied from chamber reactions of uo- Ambient particles ranthene, pyrene, triphenylene, benz[a]anthracene, and chrysene with OH and NO3 radicals at room Nitrotriphenylenes temperature and atmospheric pressure, with the PAH concentrations being controlled by their vapor pressures. -

Questions & Answers for the New Chemicals Program
Note: Effective January 19, 2016, PMNs must be submitted electronically. Learn more about the new e-PMN requirements. Questions & Answers for the New Chemicals Program (Q&A) U.S. Environmental Protection Agency Office of Pollution Prevention and Toxics Washington, DC 20460 2004 -1- TABLE OF CONTENTS Page 1. GENERAL PROGRAM INFORMATION 100. General ............................................................................................................ 1-1 101. Guidance for Completion of §5 Submission Form ......................................... 1-6 102. Inventory Searches/Bona Fides ....................................................................... 1-17 103. Chemical Identification ................................................................................... 1-22 104. Nomenclature .................................................................................................. 1-26 105. Inventory Issues ................................................................................................ 1-31 106. Review Process ............................................................................................... 1-31 107. Notice of Commencement .............................................................................. 1-33 108. User Fee .......................................................................................................... 1-35 109. Consolidated Notices ...................................................................................... 1-39 110. Joint Submissions .......................................................................................... -

Electrophilic Mercuration and Thallation of Benzene and Substituted Benzenes in Trifluoroacetic Acid Solution* (Electrophilic Substitution/Selectivity) GEORGE A
Proc. Natl. Acad. Sci. USA Vol. 74, No. 10, pp. 4121-4125, October 1977 Chemistry Electrophilic mercuration and thallation of benzene and substituted benzenes in trifluoroacetic acid solution* (electrophilic substitution/selectivity) GEORGE A. OLAH, IWAO HASHIMOTOt, AND HENRY C. LINtt Institute of Hydrocarbon Chemistry, Department of Chemistry, University of Southern California, Los Angeles, California 90007; and the Department of Chemistry, Case Western Reserve University, Cleveland, Ohio 44101 Contributed by George A. Olah, July 18, 1977 ABSTRACT The mercuration and thallation of benzene and iments at 250 and quenched the mixtures after 5 min, whereas substituted benzenes was studied with mercuric and thallic Brown and Nelson carried out the reaction for 6.5 hr. In sub- trifluoroacetate, respectively, in trifluoroacetic acid. With the in view of the importance of both the mechanistic shortest reaction time (1 sec) at 00, the relative rate of mercu- sequent work, ration of toluene compared to that of benzene was 17.5, with and practical implications of the orientation-rate correlation, the isomer distribution in toluene of: ortho, 17.4%; meta, 5.9%; Brown and McGary (7), carried out a more detailed study of and para, 76.7%. The isomer distribution in toluene varied with the mercuration reaction. They concluded: "A redetermination the reaction time, significantly more at 25° than at 00. The of the isomer distributions and relative rates indicates excellent competitive thallation of benzene and toluene with thallic tri- agreement with the linear relationship of orientation and rel- fluoroacetate in trifluoroacetic acid at 150 showed the relative ative however, that the isomer distri- rate, toluene/benzene, to be 33, with the isomer distribution in rate." They recognized, toluene of: ortho, 9.5%; meta, 5.5%; and para, 85.0%. -

Cis-Trans Isomerism 2
Isomerism Isomerism • These compounds possess the same molecular formula but differ from each other in physical or chemical properties, and are called Isomers and the phenomenon is termed Isomerism • Since isomers have the same molecular formula, the difference in their properties must be due to different modes of combination or arrangement of atoms within the molecule. There are two main types of isomerism • (i) Structural Isomerism and (ii) Stereoisomerism. Structural Isomerism • When the isomerism is simply due to difference in the arrangement of atoms within the molecule without any reference to space, the phenomenon is termed Structural Isomerism. In other words, the structural isomers are compounds that have the same molecular formula but different structural formulas. Types of Structural Isomerism 1. Chain Isomerism 2. Position Isomerism 3. Functional Isomerism 4. Metamersim 5. Tautomerism Stereoisomerism • When isomerism is caused by the different arrangements of atoms or groups in space, the phenomenon is called Stereoisomerism (Greek, Stereos = occupying space). • The stereoisomers have the same structural formulas but differ in arrangement of atoms in space. In other words, stereoisomerism is exhibited by such compounds which have identical molecular structure but different configurations. Configuration refers to the 3-dimentional arrangement of atoms that characterizes a particular compound. Types of Stereoisomerism Stereoisomerism is of two types 1. Geometrical or Cis-Trans Isomerism 2. Optical Isomerism Structural Isomerism Chain Isomerism This type of isomerism arises from she difference in the structure of carbon chain which forms the nucleus of the molecule. It is, therefore, named as Chain or Nuclear Isomerism. Chain isomers have the same molecular formula but differ in the order in which the carbon atoms are bonded to each other For example, there are known two butanes which have the same molecular formula (C4H10) but differ in the structure of the carbon chains in their molecules. -

Alkenes and Alkynes
02/21/2019 CHAPTER FOUR Alkenes and Alkynes H N O I Cl C O C O Cl F3C C Cl C Cl Efavirenz Haloprogin (antiviral, AIDS therapeutic) (antifungal, antiseptic) Chapter 4 Table of Content * Unsaturated Hydrocarbons * Introduction and hybridization * Alkenes and Alkynes * Benzene and Phenyl groups * Structure of Alkenes, cis‐trans Isomerism * Nomenclature of Alkenes and Alkynes * Configuration cis/trans, and cis/trans Isomerism * Configuration E/Z * Physical Properties of Hydrocarbons * Acid‐Base Reactions of Hydrocarbons * pka and Hybridizations 1 02/21/2019 Unsaturated Hydrocarbons • Unsaturated Hydrocarbon: A hydrocarbon that contains one or more carbon‐carbon double or triple bonds or benzene‐like rings. – Alkene: contains a carbon‐carbon double bond and has the general formula CnH2n. – Alkyne: contains a carbon‐carbon triple bond and has the general formula CnH2n‐2. Introduction Alkenes ● Hydrocarbons containing C=C ● Old name: olefins • Steroids • Hormones • Biochemical regulators 2 02/21/2019 • Alkynes – Hydrocarbons containing C≡C – Common name: acetylenes Unsaturated Hydrocarbons • Arene: benzene and its derivatives (Ch 9) 3 02/21/2019 Benzene and Phenyl Groups • We do not study benzene and its derivatives until Chapter 9. – However, we show structural formulas of compounds containing a phenyl group before that time. – The phenyl group is not reactive under any of the conditions we describe in chapters 5‐8. Structure of Alkenes • The two carbon atoms of a double bond and the four atoms bonded to them lie in a plane, with bond angles of approximately 120°. 4 02/21/2019 Structure of Alkenes • Figure 4.1 According to the orbital overlap model, a double bond consists of one bond formed by overlap of sp2 hybrid orbitals and one bond formed by overlap of parallel 2p orbitals. -

Enantioselective and Synergistic Herbicidal Activities of Common Amino Acids Against Amaranthus Tricolor and Echinochloa Crus-Galli
molecules Article Enantioselective and Synergistic Herbicidal Activities of Common Amino Acids Against Amaranthus tricolor and Echinochloa crus-galli Nawasit Chotsaeng 1,2,* , Chamroon Laosinwattana 3 and Patchanee Charoenying 1 1 Department of Chemistry, School of Science, King Mongkut’s Institute of Technology Ladkrabang, Bangkok 10520, Thailand; [email protected] 2 Integrated Applied Chemistry Research Unit, School of Science, King Mongkut’s Institute of Technology Ladkrabang, Bangkok 10520, Thailand 3 Department of Plant Production Technology, School of Agricultural Technology, King Mongkut’s Institute of Technology Ladkrabang, Bangkok 10520, Thailand; [email protected] * Correspondence: [email protected] or [email protected]; Tel.: +66-2329-8400 (ext. 6228); Fax: +66-2329-8428 Abstract: Amino acids have a wide range of biological activities, which usually rely on the stereoiso- mer presented. In this study, glycine and 21 common α-amino acids were investigated for their herbicidal property against Chinese amaranth (Amaranthus tricolor L.) and barnyard grass (Echinochloa crus-galli (L.) Beauv.). Both D- and L-isomers, as well as a racemic mixture, were tested and found that most compounds barely inhibited germination but moderately suppressed seedling growth. Various ratios of D:L-mixture were studied and synergy between enantiomers was found. For Chinese amaranth, the most toxic D:L-mixtures were at 3:7 (for glutamine), 8:2 (for methionine), and 5:5 (for tryptophan). For barnyard grass, rac-glutamine was more toxic than the pure forms; Citation: Chotsaeng, N.; however, D-tryptophan exhibited greater activity than racemate and L-isomer, indicating the sign Laosinwattana, C.; Charoenying, P. -

Highly Enantioselective Synthesis of Γ-, Δ-, and E-Chiral 1-Alkanols Via Zr-Catalyzed Asymmetric Carboalumination of Alkenes (ZACA)–Cu- Or Pd-Catalyzed Cross-Coupling
Highly enantioselective synthesis of γ-, δ-, and e-chiral 1-alkanols via Zr-catalyzed asymmetric carboalumination of alkenes (ZACA)–Cu- or Pd-catalyzed cross-coupling Shiqing Xu, Akimichi Oda, Hirofumi Kamada, and Ei-ichi Negishi1 Department of Chemistry, Purdue University, West Lafayette, IN 47907 Edited by Chi-Huey Wong, Academia Sinica, Taipei, Taiwan, and approved May 2, 2014 (received for review January 21, 2014) Despite recent advances of asymmetric synthesis, the preparation shown in Schemes 1 and 2 illustrate the versatility of ZACA of enantiomerically pure (≥99% ee) compounds remains a chal- represented by the organoaluminum functionality of the ini- lenge in modern organic chemistry. We report here a strategy tially formed ZACA products. Introduction of the OH group for a highly enantioselective (≥99% ee) and catalytic synthesis by oxidation of initially formed alkylalane intermediates in of various γ- and more-remotely chiral alcohols from terminal Scheme 2 is based on two considerations: (i) the proximity of the alkenes via Zr-catalyzed asymmetric carboalumination of alkenes OH group to a stereogenic carbon center is highly desirable for (ZACA reaction)–Cu- or Pd-catalyzed cross-coupling. ZACA–in situ lipase-catalyzed acetylation to provide ultrapure (≥99% ee) di- oxidation of tert-butyldimethylsilyl (TBS)-protected ω-alkene-1-ols functional intermediates, and (ii) the versatile OH group can R S α ω produced both ( )- and ( )- , -dioxyfunctional intermediates (3) be further transformed to a wide range of carbon groups by – ee ≥ ee in 80 88% , which were readily purified to the 99% level tosylation or iodination followed by Cu- or Pd-catalyzed cross- by lipase-catalyzed acetylation through exploitation of their high α ω coupling. -

1.5 Isomers Let’S Look at the Following 2 Molecules
1.5 Isomers Let’s look at the following 2 molecules: Note that although these 2 structures have the same atomic composition (C2H4F2), in the first molecule there is one F on each of the C atoms whereas on the second molecule both F atoms are on the same C atom. No matter how we rotate the molecule or turn it over or around, these 2 structures are fundamentally different molecules. We call these molecules structural or constitutional isomers. Definition: Molecules which have the same formula, but which have a difference in structure (in what atoms are bonded to each other) are called structural or constitutional isomers. For each of the pairs of molecular structures shown below, are the pairs of molecules isomers or just different representations of the same molecule? (Answer: These are different representations of the same structure because of free rotation around the C-Cl bonds. We can rotate the C-Cl bonds in any direction we want without creating a new structure.) (Answer: These are structural isomers because the structure on the left has 2 Br atoms on one C atom and 1 Br atom on the other. The second structure has all 3 Br atoms on one C atom. We can’t rotate or flip our way out of this one. These two structures have the same formula but different bonding and are hence structural isomers.) (Answer: Each C atom has one F atom in the structure on the left, while the structure on the right has both F atoms on the same C atom. -

1 Chapter 3: Organic Compounds: Alkanes and Cycloalkanes
Chapter 3: Organic Compounds: Alkanes and Cycloalkanes >11 million organic compounds which are classified into families according to structure and reactivity Functional Group (FG): group of atoms which are part of a large molecule that have characteristic chemical behavior. FG’s behave similarly in every molecule they are part of. The chemistry of the organic molecule is defined by the function groups it contains 1 C C Alkanes Carbon - Carbon Multiple Bonds Carbon-heteroatom single bonds basic C N C C C X X= F, Cl, Br, I amines Alkenes Alkyl Halide H C C C O C C O Alkynes alcohols ethers acidic H H H C S C C C C S C C H sulfides C C thiols (disulfides) H H Arenes Carbonyl-oxygen double bonds (carbonyls) Carbon-nitrogen multiple bonds acidic basic O O O N H C H C O C Cl imine (Schiff base) aldehyde carboxylic acid acid chloride O O O O C C N C C C C O O C C nitrile (cyano group) ketones ester anhydrides O C N amide opsin Lys-NH2 + Lys- opsin H O H N rhodopsin H 2 Alkanes and Alkane Isomers Alkanes: organic compounds with only C-C and C-H single (s) bonds. general formula for alkanes: CnH(2n+2) Saturated hydrocarbons Hydrocarbons: contains only carbon and hydrogen Saturated" contains only single bonds Isomers: compounds with the same chemical formula, but different arrangement of atoms Constitutional isomer: have different connectivities (not limited to alkanes) C H O C4H10 C5H12 2 6 O OH butanol diethyl ether straight-chain or normal hydrocarbons branched hydrocarbons n-butane n-pentane Systematic Nomenclature (IUPAC System) Prefix-Parent-Suffix -

2.4 Constitutional Isomers and Nomenclature 57
02_BRCLoudon_pgs4-4.qxd 11/26/08 8:36 AM Page 57 2.4 CONSTITUTIONAL ISOMERS AND NOMENCLATURE 57 H CH3CH2 H H CH(CH3)2 H 2-methylhexane anti conformation about C3–C4 bond Remember that Newman projections are used to examine conformations about a particular bond. If we want to examine the conformations about several different bonds, we must draw a different set of Newman projections for each bond. PROBLEMS 2.3 (a) Draw a Newman projection for each conformation about the C2 C3 bond of isopentane, a compound containing a branched carbon chain. L 2 3 H3C CH CH2 CH3 L L L "CH3 isopentane Show both staggered and eclipsed conformations. (b) Sketch a curve of potential energy versus dihedral angle for isopentane, similar to that of butane in Fig. 2.5. Label each energy maximum and minimum with one of the conforma- tions you drew in part (a). (c) Which conformations are likely to be present in greatest amount in a sample of isopentane? Explain. 2.4 Repeat the analysis in Problem 2.3 for either one of the terminal bonds of butane. 2.4 CONSTITUTIONAL ISOMERS AND NOMENCLATURE A. Isomers When a carbon atom in an alkane is bound to more than two other carbon atoms, a branch in the carbon chain occurs at that position. The smallest branched alkane has four carbon atoms. As a result, there are two four-carbon alkanes; one is butane, and the other is isobutane. H3C H3C CH2 CH2 CH3 $CH CH3 L L L L butane H3C) bp 0.5° - isobutane bp 11.7° - These are different compounds with different properties. -

Isomers and Coordination Geometries Chapter 9
Coordination Chemistry II: Isomers and Coordination Geometries Chapter 9 Monday, November 16, 2015 A Real World Example of Stuff from Class! Isomerism Coordination complexes often have a variety of isomeric forms Structural Isomers Molecules with the same numbers of the same atoms, but in different arrangements. Isomers generally have distinct physical and chemical properties. One isomer may be a medicine while another is a poison. Type 1: Structural isomers differ in how the atoms are connected. As a result, they have different chemical formulas. e.g., C3H8O 1-propanol 2-propanol methoxyethane m.p. -127°C m.p. -89°C m.p. -139°C b.p. 97°C b.p. 83°C b.p. 8°C Structural Isomers Structural (or constitutional) isomers are molecules with the same kind and number of atoms but with different bond arrangements In coordination complexes there are four types of structural isomers: • hydrate (solvent) isomerism occurs when water (or another solvent) can appear within the primary or secondary coordination sphere of a metal ion Cr H O Cl CrCl H O Cl H O 2 6 3 2 5 2 2 violet crystals blue-green crystals CrCl H O Cl 2H O CrCl H O 3H O 2 2 4 2 3 2 3 2 dark green crystals yellow-green crystals • ionization isomers afford different anions and cations in solution Co NH SO NO Co NH NO SO 3 5 4 3 3 5 3 4 Structural Isomers In coordination complexes there are four types of structural isomers • coordination isomerism occurs when ligands can be distributed differently between two or more metals • linkage isomerism occurs when a ligand can bind in different ways to a metal Stereoisomers The second major class of isomer.