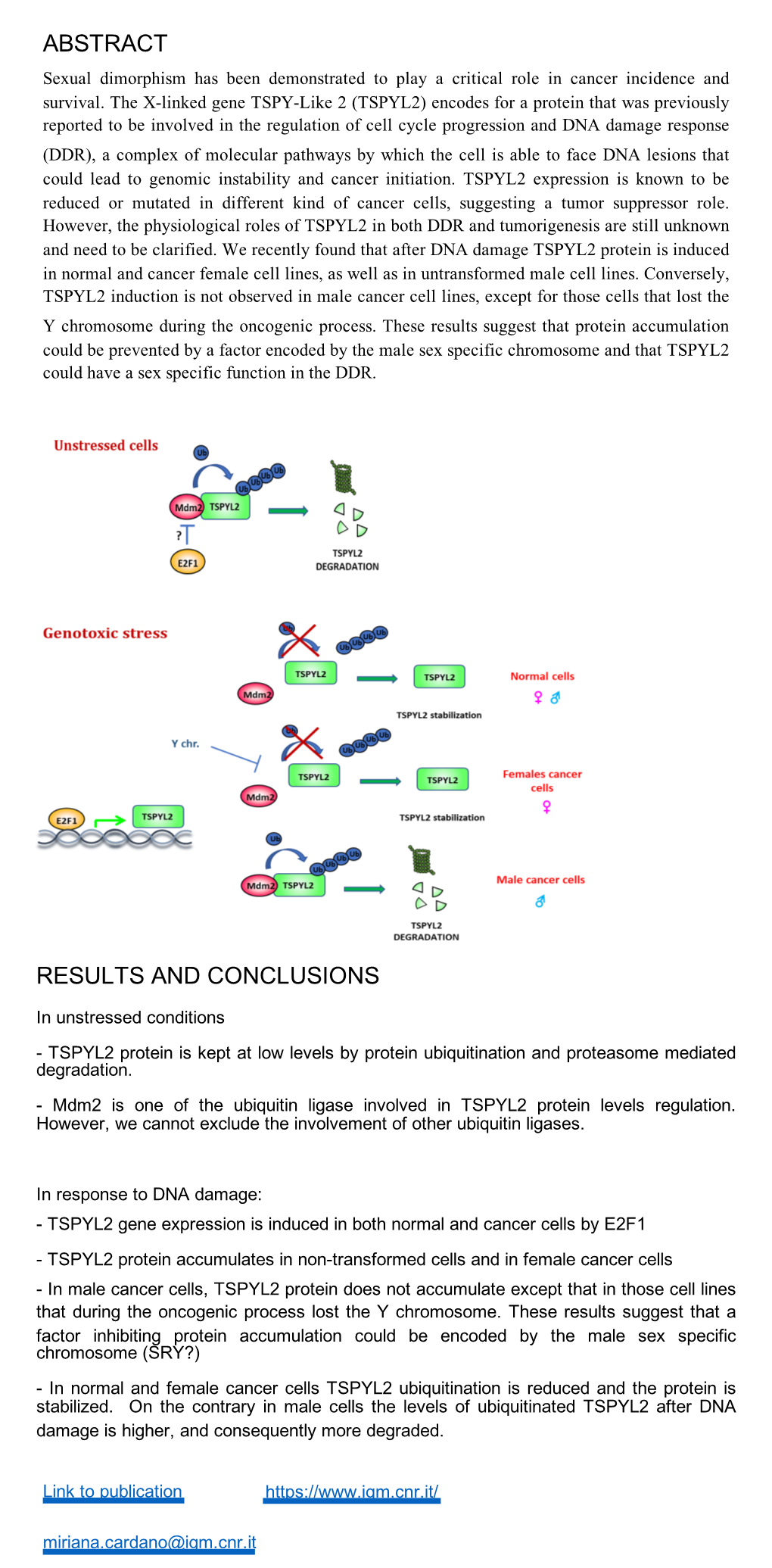
TSPYL2 Is a Novel Regulator of SIRT1 and P300 Activity in Response to DNA Damage
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Escape from X Chromosome Inactivation Is an Intrinsic Property of the Jarid1c Locus
Escape from X chromosome inactivation is an intrinsic property of the Jarid1c locus Nan Lia,b and Laura Carrela,1 aDepartment of Biochemistry and Molecular Biology and bIntercollege Graduate Program in Genetics, Pennsylvania State College of Medicine, Hershey, PA 17033 Edited by Stanley M. Gartler, University of Washington, Seattle, WA, and approved September 23, 2008 (received for review August 8, 2008) Although most genes on one X chromosome in mammalian fe- Sequences on the X are hypothesized to propagate XCI (12) and males are silenced by X inactivation, some ‘‘escape’’ X inactivation to be depleted at escape genes (8, 9). LINE-1 repeats fit such and are expressed from both active and inactive Xs. How these predictions, particularly on the human X (8–10). Distinct dis- escape genes are transcribed from a largely inactivated chromo- tributions of other repeats classify some mouse X genes (5). some is not fully understood, but underlying genomic sequences X-linked transgenes also test the role of genomic sequences in are likely involved. We developed a transgene approach to ask escape gene expression. Most transgenes are X-inactivated, whether an escape locus is autonomous or is instead influenced by although a number escape XCI (e.g., refs. 13 and 14). Such X chromosome location. Two BACs carrying the mouse Jarid1c gene transgene studies indicate that, in addition to CTCF (6), locus and adjacent X-inactivated transcripts were randomly integrated control regions and matrix attachment sites are also not suffi- into mouse XX embryonic stem cells. Four lines with single-copy, cient to escape XCI (15, 16). -

Environmental Influences on Endothelial Gene Expression
ENDOTHELIAL CELL GENE EXPRESSION John Matthew Jeff Herbert Supervisors: Prof. Roy Bicknell and Dr. Victoria Heath PhD thesis University of Birmingham August 2012 University of Birmingham Research Archive e-theses repository This unpublished thesis/dissertation is copyright of the author and/or third parties. The intellectual property rights of the author or third parties in respect of this work are as defined by The Copyright Designs and Patents Act 1988 or as modified by any successor legislation. Any use made of information contained in this thesis/dissertation must be in accordance with that legislation and must be properly acknowledged. Further distribution or reproduction in any format is prohibited without the permission of the copyright holder. ABSTRACT Tumour angiogenesis is a vital process in the pathology of tumour development and metastasis. Targeting markers of tumour endothelium provide a means of targeted destruction of a tumours oxygen and nutrient supply via destruction of tumour vasculature, which in turn ultimately leads to beneficial consequences to patients. Although current anti -angiogenic and vascular targeting strategies help patients, more potently in combination with chemo therapy, there is still a need for more tumour endothelial marker discoveries as current treatments have cardiovascular and other side effects. For the first time, the analyses of in-vivo biotinylation of an embryonic system is performed to obtain putative vascular targets. Also for the first time, deep sequencing is applied to freshly isolated tumour and normal endothelial cells from lung, colon and bladder tissues for the identification of pan-vascular-targets. Integration of the proteomic, deep sequencing, public cDNA libraries and microarrays, delivers 5,892 putative vascular targets to the science community. -

Open Data for Differential Network Analysis in Glioma
International Journal of Molecular Sciences Article Open Data for Differential Network Analysis in Glioma , Claire Jean-Quartier * y , Fleur Jeanquartier y and Andreas Holzinger Holzinger Group HCI-KDD, Institute for Medical Informatics, Statistics and Documentation, Medical University Graz, Auenbruggerplatz 2/V, 8036 Graz, Austria; [email protected] (F.J.); [email protected] (A.H.) * Correspondence: [email protected] These authors contributed equally to this work. y Received: 27 October 2019; Accepted: 3 January 2020; Published: 15 January 2020 Abstract: The complexity of cancer diseases demands bioinformatic techniques and translational research based on big data and personalized medicine. Open data enables researchers to accelerate cancer studies, save resources and foster collaboration. Several tools and programming approaches are available for analyzing data, including annotation, clustering, comparison and extrapolation, merging, enrichment, functional association and statistics. We exploit openly available data via cancer gene expression analysis, we apply refinement as well as enrichment analysis via gene ontology and conclude with graph-based visualization of involved protein interaction networks as a basis for signaling. The different databases allowed for the construction of huge networks or specified ones consisting of high-confidence interactions only. Several genes associated to glioma were isolated via a network analysis from top hub nodes as well as from an outlier analysis. The latter approach highlights a mitogen-activated protein kinase next to a member of histondeacetylases and a protein phosphatase as genes uncommonly associated with glioma. Cluster analysis from top hub nodes lists several identified glioma-associated gene products to function within protein complexes, including epidermal growth factors as well as cell cycle proteins or RAS proto-oncogenes. -

A High-Throughput Approach to Uncover Novel Roles of APOBEC2, a Functional Orphan of the AID/APOBEC Family
Rockefeller University Digital Commons @ RU Student Theses and Dissertations 2018 A High-Throughput Approach to Uncover Novel Roles of APOBEC2, a Functional Orphan of the AID/APOBEC Family Linda Molla Follow this and additional works at: https://digitalcommons.rockefeller.edu/ student_theses_and_dissertations Part of the Life Sciences Commons A HIGH-THROUGHPUT APPROACH TO UNCOVER NOVEL ROLES OF APOBEC2, A FUNCTIONAL ORPHAN OF THE AID/APOBEC FAMILY A Thesis Presented to the Faculty of The Rockefeller University in Partial Fulfillment of the Requirements for the degree of Doctor of Philosophy by Linda Molla June 2018 © Copyright by Linda Molla 2018 A HIGH-THROUGHPUT APPROACH TO UNCOVER NOVEL ROLES OF APOBEC2, A FUNCTIONAL ORPHAN OF THE AID/APOBEC FAMILY Linda Molla, Ph.D. The Rockefeller University 2018 APOBEC2 is a member of the AID/APOBEC cytidine deaminase family of proteins. Unlike most of AID/APOBEC, however, APOBEC2’s function remains elusive. Previous research has implicated APOBEC2 in diverse organisms and cellular processes such as muscle biology (in Mus musculus), regeneration (in Danio rerio), and development (in Xenopus laevis). APOBEC2 has also been implicated in cancer. However the enzymatic activity, substrate or physiological target(s) of APOBEC2 are unknown. For this thesis, I have combined Next Generation Sequencing (NGS) techniques with state-of-the-art molecular biology to determine the physiological targets of APOBEC2. Using a cell culture muscle differentiation system, and RNA sequencing (RNA-Seq) by polyA capture, I demonstrated that unlike the AID/APOBEC family member APOBEC1, APOBEC2 is not an RNA editor. Using the same system combined with enhanced Reduced Representation Bisulfite Sequencing (eRRBS) analyses I showed that, unlike the AID/APOBEC family member AID, APOBEC2 does not act as a 5-methyl-C deaminase. -

Volume 18 - Number 1 January 2014
Atlas of Genetics and Cytogenetics in Oncology and Haematology OPEN ACCESS JOURNAL INIST -CNRS Volume 18 - Number 1 January 2014 The PDF version of the Atlas of Genetics and Cytogenetics in Oncology and Haematology is a reissue of the original articles published in collaboration with the Institute for Scientific and Technical Information (INstitut de l’Information Scientifique et Technique - INIST) of the French National Center for Scientific Research (CNRS) on its electronic publishing platform I-Revues. Online and PDF versions of the Atlas of Genetics and Cytogenetics in Oncology and Haematology are hosted by INIST-CNRS. Atlas of Genetics and Cytogenetics in Oncology and Haematology OPEN ACCESS JOURNAL INIST -CNRS Scope The Atlas of Genetics and Cytogenetics in Oncology and Haematology is a peer reviewed on-line journal in open access, devoted to genes, cytogenetics, and clinical entities in cancer, and cancer-prone diseases. It presents structured review articles (“cards”) on genes, leukaemias, solid tumours, cancer-prone diseases, and also more traditional review articles (“deep insights”) on the above subjects and on surrounding topics. It also present case reports in hematology and educational items in the various related topics for students in Medicine and in Sciences. Editorial correspondance Jean-Loup Huret Genetics, Department of Medical Information, University Hospital F-86021 Poitiers, France tel +33 5 49 44 45 46 or +33 5 49 45 47 67 [email protected] or [email protected] Staff Mohammad Ahmad, Mélanie Arsaban, Marie-Christine Jacquemot-Perbal, Vanessa Le Berre, Anne Malo, Carol Moreau, Catherine Morel-Pair, Laurent Rassinoux, Alain Zasadzinski. Philippe Dessen is the Database Director, and Alain Bernheim the Chairman of the on-line version (Gustave Roussy Institute – Villejuif – France). -

CREB-Dependent Transcription in Astrocytes: Signalling Pathways, Gene Profiles and Neuroprotective Role in Brain Injury
CREB-dependent transcription in astrocytes: signalling pathways, gene profiles and neuroprotective role in brain injury. Tesis doctoral Luis Pardo Fernández Bellaterra, Septiembre 2015 Instituto de Neurociencias Departamento de Bioquímica i Biologia Molecular Unidad de Bioquímica y Biologia Molecular Facultad de Medicina CREB-dependent transcription in astrocytes: signalling pathways, gene profiles and neuroprotective role in brain injury. Memoria del trabajo experimental para optar al grado de doctor, correspondiente al Programa de Doctorado en Neurociencias del Instituto de Neurociencias de la Universidad Autónoma de Barcelona, llevado a cabo por Luis Pardo Fernández bajo la dirección de la Dra. Elena Galea Rodríguez de Velasco y la Dra. Roser Masgrau Juanola, en el Instituto de Neurociencias de la Universidad Autónoma de Barcelona. Doctorando Directoras de tesis Luis Pardo Fernández Dra. Elena Galea Dra. Roser Masgrau In memoriam María Dolores Álvarez Durán Abuela, eres la culpable de que haya decidido recorrer el camino de la ciencia. Que estas líneas ayuden a conservar tu recuerdo. A mis padres y hermanos, A Meri INDEX I Summary 1 II Introduction 3 1 Astrocytes: physiology and pathology 5 1.1 Anatomical organization 6 1.2 Origins and heterogeneity 6 1.3 Astrocyte functions 8 1.3.1 Developmental functions 8 1.3.2 Neurovascular functions 9 1.3.3 Metabolic support 11 1.3.4 Homeostatic functions 13 1.3.5 Antioxidant functions 15 1.3.6 Signalling functions 15 1.4 Astrocytes in brain pathology 20 1.5 Reactive astrogliosis 22 2 The transcription -

Functional Repertoire of EV-Associated Mirna Profiles After Lipoprotein Depletion Via Ultracentrifugation and Size Exclusion
www.nature.com/scientificreports OPEN Functional repertoire of EV‑associated miRNA profles after lipoprotein depletion via ultracentrifugation and size exclusion chromatography from autologous blood products Alexander Otahal1, Olga Kuten‑Pella2, Karina Kramer1, Markus Neubauer1, Zsombor Lacza3, Stefan Nehrer1 & Andrea De Luna1* Cartilage breakdown, infammation and pain are hallmark symptoms of osteoarthritis, and autologous blood products such as citrate‑anticoagulated platelet‑rich plasma (CPRP) or hyperacute serum (hypACT) have been developed as a regenerative approach to rebuild cartilage, inhibit infammation and reduce pain. However, mechanisms of action of these blood derivatives are still not fully understood, in part due to the large number of components present in these medical products. In addition, the discovery of extracellular vesicles (EVs) and their involvement in intercellular communication mediated by cargo molecules like microRNAs (miRNAs) opened up a whole new level of complexity in understanding blood products. In this study we focused on the development of an isolation protocol for EVs from CPRP and hypACT that can also deplete lipoproteins, which are often co‑isolated in EV research due to shared physical properties. Several isolation methods were compared in terms of particle yield from CPRP and hypACT. To gain insights into the functional repertoire conveyed via EV‑associated miRNAs, we performed functional enrichment analysis and identifed NFκB signaling strongly targeted by CPRP EV miRNAs, whereas hypACT EV miRNAs afect IL6‑ and TGFβ/SMAD signaling. Autologous blood derived products have already been used for more than a decade and increasingly gain popu- larity in regenerative medicine. A main application is the intra-articular injection into afected joints of osteo- arthritis (OA) patients1, besides other treatments of muskuloskeletal conditions such as epicondylitis or rotator cuf injuries2,3. -

Museum of London Report on the DNA Analyses of Four Roman Individuals Supplementary Information
Museum of London Report on the DNA Analyses of Four Roman Individuals Supplementary Information Katherine Eaton1, Ana T. Duggan1, Alison Devault2, and Hendrik Poinar1 1McMaster Ancient DNA Centre, Department of Anthropology, McMaster University, Hamilton, ON, Canada 2MYcroarray, Ann Arbor, MI, USA November 27, 2015 1 Contents 1 Sample Information3 2 Pre-PCR Labwork3 2.1 Subsampling, Demineralization, Protein Digestion...................3 2.2 Extraction, Concentration, Purification.........................4 3 Library Preparation and Indexing4 4 In-Solution Bait Design4 4.1 Human Mitochondrial Baits...............................4 4.2 Hair and Eye Color....................................4 4.3 Sex Identification.....................................5 5 Targeted Enrichment8 5.1 Human Mitochondria, HIrisPlex, Sex..........................8 6 Sequencing 9 6.1 Mitochondrial Haplotype.................................9 6.2 Geographic Distribution of Haplogroups........................ 16 6.3 HIrisPlex Genotype.................................... 17 6.4 Sex Estimation...................................... 18 7 Summary 20 2 1 Sample Information The four individuals examined in this study are permanently stored at the Museum of London. Osteobiographical data and excavation context for these individuals are summarized in Table1. Table 1: Summary of Skeletal Characteristics and Excavation Context Individual ID Location Time Period Skeletal Sex Age HR79 SK311 T13 Harper Road, Southwark 50-70 AD Female >45 Years MNL88 SK37 T16 65-73 Mansell Street 180-400 AD Male >45 Years LTU03 SK385 T17 Lant Street, Southwark 300-400 AD Female 14 Years LOW88 SK695.5 T18 London Wall 125-200 AD Male 36-45 Years Subsampling and all subsequent labwork was conducted at the McMaster Ancient DNA Centre. All pre-PCR work, including subsampling, extraction, and library prep, was conducted in dedicated ancient DNA clean rooms that have not been exposed to high-copy modern DNA or high copy PCR products. -

Next Generation Mapping Reveals Novel Large Genomic Rearrangements in Prostate Cancer
www.impactjournals.com/oncotarget/ Oncotarget, 2017, Vol. 8, (No. 14), pp: 23588-23602 Research Paper Next generation mapping reveals novel large genomic rearrangements in prostate cancer Weerachai Jaratlerdsiri1,*, Eva K.F. Chan1,2,*, Desiree C. Petersen1,2,*, Claire Yang3, Peter I. Croucher2,4,5, M.S. Riana Bornman6, Palak Sheth3, Vanessa M. Hayes1,2,6,7 1Laboratory for Human Comparative and Prostate Cancer Genomics, Genomics and Epigenetics Division, Garvan Institute of Medical Research, Darlinghurst, Australia 2St Vincent’s Clinical School, University of New South Wales, Randwick, Australia 3Bionano Genomics Inc., San Diego, California, USA 4Bone Biology Division, Garvan Institute of Medical Research, Darlinghurst, Australia 5School of Biotechnology and Biomolecular Sciences, University of New South Wales, Randwick, Australia 6School of Health Systems and Public Health, University of Pretoria, Pretoria, South Africa 7Central Clinical School, University of Sydney, Camperdown, Australia *These authors contributed equally to this work Correspondence to: Vanessa M. Hayes, email: [email protected] Keywords: prostate cancer, structural genomic rearrangements, next generation mapping, next generation sequencing Received: August 15, 2016 Accepted: February 15, 2017 Published: March 01, 2017 ABSTRACT Complex genomic rearrangements are common molecular events driving prostate carcinogenesis. Clinical significance, however, has yet to be fully elucidated. Detecting the full range and subtypes of large structural variants (SVs), greater than -

Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress
University of Pennsylvania ScholarlyCommons Publicly Accessible Penn Dissertations Fall 2010 Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress Renuka Nayak University of Pennsylvania, [email protected] Follow this and additional works at: https://repository.upenn.edu/edissertations Part of the Computational Biology Commons, and the Genomics Commons Recommended Citation Nayak, Renuka, "Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress" (2010). Publicly Accessible Penn Dissertations. 1559. https://repository.upenn.edu/edissertations/1559 This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/1559 For more information, please contact [email protected]. Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress Abstract Genes interact in networks to orchestrate cellular processes. Here, we used coexpression networks based on natural variation in gene expression to study the functions and interactions of human genes. We asked how these networks change in response to stress. First, we studied human coexpression networks at baseline. We constructed networks by identifying correlations in expression levels of 8.9 million gene pairs in immortalized B cells from 295 individuals comprising three independent samples. The resulting networks allowed us to infer interactions between biological processes. We used the network to predict the functions of poorly-characterized human genes, and provided some experimental support. Examining genes implicated in disease, we found that IFIH1, a diabetes susceptibility gene, interacts with YES1, which affects glucose transport. Genes predisposing to the same diseases are clustered non-randomly in the network, suggesting that the network may be used to identify candidate genes that influence disease susceptibility. -

Distinct Transcriptomes Define Rostral and Caudal 5Ht Neurons
DISTINCT TRANSCRIPTOMES DEFINE ROSTRAL AND CAUDAL 5HT NEURONS by CHRISTI JANE WYLIE Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy Dissertation Advisor: Dr. Evan S. Deneris Department of Neurosciences CASE WESTERN RESERVE UNIVERSITY May, 2010 CASE WESTERN RESERVE UNIVERSITY SCHOOL OF GRADUATE STUDIES We hereby approve the thesis/dissertation of ______________________________________________________ candidate for the ________________________________degree *. (signed)_______________________________________________ (chair of the committee) ________________________________________________ ________________________________________________ ________________________________________________ ________________________________________________ ________________________________________________ (date) _______________________ *We also certify that written approval has been obtained for any proprietary material contained therein. TABLE OF CONTENTS TABLE OF CONTENTS ....................................................................................... iii LIST OF TABLES AND FIGURES ........................................................................ v ABSTRACT ..........................................................................................................vii CHAPTER 1 INTRODUCTION ............................................................................................... 1 I. Serotonin (5-hydroxytryptamine, 5HT) ....................................................... 1 A. Discovery.............................................................................................. -

Genomic Analysis Reveals Selection Signatures of Yucatan Miniature Pig on Its X Chromosome During Domestication
bioRxiv preprint doi: https://doi.org/10.1101/596387; this version posted April 4, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Genomic analysis reveals selection signatures of Yucatan miniature pig on its X chromosome during domestication Wei Zhang†, Yuanlang Wang†, Min Yang, Xudong Wu, Xiaodong Zhang, Yueyun Ding* and Zongjun Yin* College of Animal Science and Technology, Anhui Agricultural University, Hefei 230036, P. R. China †These authors contributed equally to this work. 1 bioRxiv preprint doi: https://doi.org/10.1101/596387; this version posted April 4, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Running head: Selection signatures of Yucatan pig Keywords: Yucatan miniature pig, genome variation, selection signature, odor, X chromosome. ∗Corresponding author: Zongjun Yin and Yueyun Ding Office mailing address: College of Animal Science and Technology, Anhui Agricultural University, 130, Changjiang West Road, Hefei, 230036, P. R. China. Phone number: +86-0551-65787303 E-mail: [email protected]; [email protected] Abstract Yucatan miniature pig (YMP), a naturally small breed, has been domesticated in the hot and arid Yucatan Peninsula for a long time. However, its selection signatures on the X chromosome remain poorly understood.