The Prairie Peninsula (Physiographic Area 31) Partners in Flight Bird Conservation Plan for the Prairie Peninsula (Physiographic Area 31)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Phase I Avian Risk Assessment
PHASE I AVIAN RISK ASSESSMENT Garden Peninsula Wind Energy Project Delta County, Michigan Report Prepared for: Heritage Sustainable Energy October 2007 Report Prepared by: Paul Kerlinger, Ph.D. John Guarnaccia Curry & Kerlinger, L.L.C. P.O. Box 453 Cape May Point, NJ 08212 (609) 884-2842, fax 884-4569 [email protected] [email protected] Garden Peninsula Wind Energy Project, Delta County, MI Phase I Avian Risk Assessment Garden Peninsula Wind Energy Project Delta County, Michigan Executive Summary Heritage Sustainable Energy is proposing a utility-scale wind-power project of moderate size for the Garden Peninsula on the Upper Peninsula of Michigan in Delta County. This peninsula separates northern Lake Michigan from Big Bay de Noc. The number of wind turbines is as yet undetermined, but a leasehold map provided to Curry & Kerlinger indicates that turbines would be constructed on private lands (i.e., not in the Lake Superior State Forest) in mainly agricultural areas on the western side of the peninsula, and possibly on Little Summer Island. For the purpose of analysis, we are assuming wind turbines with a nameplate capacity of 2.0 MW. The turbine towers would likely be about 78.0 meters (256 feet) tall and have rotors of about 39.0 m (128 feet) long. With the rotor tip in the 12 o’clock position, the wind turbines would reach a maximum height of about 118.0 m (387 feet) above ground level (AGL). When in the 6 o’clock position, rotor tips would be about 38.0 m (125 feet) AGL. However, larger turbines with nameplate capacities (up to 2.5 MW and more) reaching to 152.5 m (500 feet) are may be used. -

Geomorphic Classification of Rivers
9.36 Geomorphic Classification of Rivers JM Buffington, U.S. Forest Service, Boise, ID, USA DR Montgomery, University of Washington, Seattle, WA, USA Published by Elsevier Inc. 9.36.1 Introduction 730 9.36.2 Purpose of Classification 730 9.36.3 Types of Channel Classification 731 9.36.3.1 Stream Order 731 9.36.3.2 Process Domains 732 9.36.3.3 Channel Pattern 732 9.36.3.4 Channel–Floodplain Interactions 735 9.36.3.5 Bed Material and Mobility 737 9.36.3.6 Channel Units 739 9.36.3.7 Hierarchical Classifications 739 9.36.3.8 Statistical Classifications 745 9.36.4 Use and Compatibility of Channel Classifications 745 9.36.5 The Rise and Fall of Classifications: Why Are Some Channel Classifications More Used Than Others? 747 9.36.6 Future Needs and Directions 753 9.36.6.1 Standardization and Sample Size 753 9.36.6.2 Remote Sensing 754 9.36.7 Conclusion 755 Acknowledgements 756 References 756 Appendix 762 9.36.1 Introduction 9.36.2 Purpose of Classification Over the last several decades, environmental legislation and a A basic tenet in geomorphology is that ‘form implies process.’As growing awareness of historical human disturbance to rivers such, numerous geomorphic classifications have been de- worldwide (Schumm, 1977; Collins et al., 2003; Surian and veloped for landscapes (Davis, 1899), hillslopes (Varnes, 1958), Rinaldi, 2003; Nilsson et al., 2005; Chin, 2006; Walter and and rivers (Section 9.36.3). The form–process paradigm is a Merritts, 2008) have fostered unprecedented collaboration potentially powerful tool for conducting quantitative geo- among scientists, land managers, and stakeholders to better morphic investigations. -

Inner Sound of Stroma, Pentland Firth (Scotland, UK)
3rd International Conference on Ocean Energy, 6 October, Bilbao An Operational Hydrodynamic Model of a key tidal-energy site: Inner Sound of Stroma, Pentland Firth (Scotland, UK) M.C. Easton 1, D.K. Woolf 1, and S. Pans 2 1 Environmental Research Institute North Highland College, UHI Millenium Institute Thurso, Caithness, KW14 7JD, Scotland Email: [email protected] 2 DHI Representative UK (Scotland) DHI Agern Allé 5, DK-2970 Hørsholm, Denmark E-mail: [email protected] Abstract mutual influence of tidal ranges and phases between North Atlantic to the West and the North Sea to the As a result of significant progress towards the East act to setup strong currents [2] where spring tides delivery of tidal-stream power the industry is commonly exceed 5 ms-1 [3-4]. moving swiftly towards the deployment of pre- Seeking to capitalise on these unique conditions and commercial arrays. Meanwhile important sites, accelerate the delivery of large-scale tidal-stream such as the Pentland Firth, are being made available energy, in March 2010 the organisation controlling the for development. It is now crucial that we consider seabed in the United Kingdom, the Crown Estate, how the installation of tidal-stream devices will awarded leases for up to 600 MW of installed tidal interact with their host environment. Surveying energy capacity within the Pentland Firth-Orkney and modelling of the marine system prior to region [5]. An additional 200 MW is expected to be development is a prerequisite to identifying any awarded later in 2010 [6]. This announcement of the significant changes associated with tidal energy first large-scale deployment of tidal energy devices deployment. -

The Gulf of Mexico Workshop on International Research, March 29–30, 2017, Houston, Texas
OCS Study BOEM 2019-045 Proceedings: The Gulf of Mexico Workshop on International Research, March 29–30, 2017, Houston, Texas U.S. Department of the Interior Bureau of Ocean Energy Management Gulf of Mexico OCS Region OCS Study BOEM 2019-045 Proceedings: The Gulf of Mexico Workshop on International Research, March 29–30, 2017, Houston, Texas Editors Larry McKinney, Mark Besonen, Kim Withers Prepared under BOEM Contract M16AC00026 by Harte Research Institute for Gulf of Mexico Studies Texas A&M University–Corpus Christi 6300 Ocean Drive Corpus Christi, TX 78412 Published by U.S. Department of the Interior New Orleans, LA Bureau of Ocean Energy Management July 2019 Gulf of Mexico OCS Region DISCLAIMER Study collaboration and funding were provided by the US Department of the Interior, Bureau of Ocean Energy Management (BOEM), Environmental Studies Program, Washington, DC, under Agreement Number M16AC00026. This report has been technically reviewed by BOEM, and it has been approved for publication. The views and conclusions contained in this document are those of the authors and should not be interpreted as representing the opinions or policies of the US Government, nor does mention of trade names or commercial products constitute endorsement or recommendation for use. REPORT AVAILABILITY To download a PDF file of this report, go to the US Department of the Interior, Bureau of Ocean Energy Management website at https://www.boem.gov/Environmental-Studies-EnvData/, click on the link for the Environmental Studies Program Information System (ESPIS), and search on 2019-045. CITATION McKinney LD, Besonen M, Withers K (editors) (Harte Research Institute for Gulf of Mexico Studies, Corpus Christi, Texas). -

Sand Dunes Computer Animations and Paper Models by Tau Rho Alpha*, John P
Go Home U.S. DEPARTMENT OF THE INTERIOR U.S. GEOLOGICAL SURVEY Sand Dunes Computer animations and paper models By Tau Rho Alpha*, John P. Galloway*, and Scott W. Starratt* Open-file Report 98-131-A - This report is preliminary and has not been reviewed for conformity with U.S. Geological Survey editorial standards. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Although this program has been used by the U.S. Geological Survey, no warranty, expressed or implied, is made by the USGS as to the accuracy and functioning of the program and related program material, nor shall the fact of distribution constitute any such warranty, and no responsibility is assumed by the USGS in connection therewith. * U.S. Geological Survey Menlo Park, CA 94025 Comments encouraged tralpha @ omega? .wr.usgs .gov [email protected] [email protected] (gobackward) <j (goforward) Description of Report This report illustrates, through computer animations and paper models, why sand dunes can develop different forms. By studying the animations and the paper models, students will better understand the evolution of sand dunes, Included in the paper and diskette versions of this report are templates for making a paper models, instructions for there assembly, and a discussion of development of different forms of sand dunes. In addition, the diskette version includes animations of how different sand dunes develop. Many people provided help and encouragement in the development of this HyperCard stack, particularly David M. Rubin, Maura Hogan and Sue Priest. -

Part 629 – Glossary of Landform and Geologic Terms
Title 430 – National Soil Survey Handbook Part 629 – Glossary of Landform and Geologic Terms Subpart A – General Information 629.0 Definition and Purpose This glossary provides the NCSS soil survey program, soil scientists, and natural resource specialists with landform, geologic, and related terms and their definitions to— (1) Improve soil landscape description with a standard, single source landform and geologic glossary. (2) Enhance geomorphic content and clarity of soil map unit descriptions by use of accurate, defined terms. (3) Establish consistent geomorphic term usage in soil science and the National Cooperative Soil Survey (NCSS). (4) Provide standard geomorphic definitions for databases and soil survey technical publications. (5) Train soil scientists and related professionals in soils as landscape and geomorphic entities. 629.1 Responsibilities This glossary serves as the official NCSS reference for landform, geologic, and related terms. The staff of the National Soil Survey Center, located in Lincoln, NE, is responsible for maintaining and updating this glossary. Soil Science Division staff and NCSS participants are encouraged to propose additions and changes to the glossary for use in pedon descriptions, soil map unit descriptions, and soil survey publications. The Glossary of Geology (GG, 2005) serves as a major source for many glossary terms. The American Geologic Institute (AGI) granted the USDA Natural Resources Conservation Service (formerly the Soil Conservation Service) permission (in letters dated September 11, 1985, and September 22, 1993) to use existing definitions. Sources of, and modifications to, original definitions are explained immediately below. 629.2 Definitions A. Reference Codes Sources from which definitions were taken, whole or in part, are identified by a code (e.g., GG) following each definition. -

Chapter 10. a First Look at Nunatsiavut Kangidualuk ('Fjord') Ecosystems
Chapter 10. A first look at Nunatsiavut Kangidualuk (‘fjord’) ecosystems Lead author Tanya Brown1,2,3, Ken Reimer3, Tom Sheldon4, Trevor Bell5 1Department of Biochemistry and Microbiology, University of Victoria, Victoria, BC; 2Institute of Ocean Sciences, Fisheries and Oceans Canada, Sidney, BC; 3Environmental Sciences Group, Royal Military College of Canada, Kingston, ON; 4Nunatsiavut Government, Nain, NF; 5Memorial University of Newfoundland, St. John’s, NF Contributing authors S. Bentley, R. Pienitz, M. Gosselin, M. Blais, M. Carpenter, E. Estrada, T. Richerol, E. Kahlmeyer, S. Luque, B. Sjare, A. Fisk, S. Iverson Abstract Long marine inlets that are classified as either fjords or fjards indent the Labrador coast. Along the moun- tainous north coast a classic fjord landscape dominates, with deep (up to ~300 m) muddy basins separated by rocky sills and flanked by high (up to 1,000 m), steep sidewalls. In contrast, the fjards of the central and southern coast are generally shallow (150 m), irregularly shaped inlets with gently sloping sidewalls and large intertidal zones. These fjords and fjards are important feeding grounds for marine mammals and seabirds and are commonly used by Inuit for hunting and travel. Despite their ecological and socio-cultural importance, these marine ecosystems are largely understudied. The Nunatsiavut Nuluak project has, as a primary goal, to undertake baseline inventories and comparative assessments of representative marine ecosystems in Labrador, including benthic and pelagic community composition, distribution and abun- dance, fjord processes, and oceanographic conditions. A case study demonstrating ecosystem resilience to anthropogenic disturbance is examined. This information provides a foundation for further research and monitoring as climate change and anthropogenic pressures alter recent baselines. -
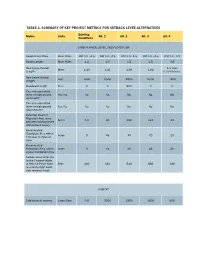
Table 2. Summary of Key Project Metrics for Setback Levee Alternatives
TABLE 2. SUMMARY OF KEY PROJECT METRICS FOR SETBACK LEVEE ALTERNATIVES Existing Metric Units Alt. 1 Alt. 2 Alt. 3 Alt. 4 Conditions CARBON RIVER, LEVEE, AND FLOODPLAIN Reach River Miles River Miles RM 3.0 - 4.5 RM 3.0 - 4.5 RM 3.0 - 4.5 RM 3.0 - 4.5 RM 3.0 - 3.9 Reach Length River Miles 1.5 1.5 1.5 1.5 0.9 New Levee Overall 1.1, plus Miles 1.43 1.61 1.69 1.84 Length 0.3 stub levee New Levee Overall Feet 7600 8500 8900 9700 7400 Length Floodwall Length Feet 0 0 500 0 0 Ties into accredited levee or high ground Yes/No No No No No No upstream? Ties into accredited levee or high ground Yes/No No No No No No downstream? Potential Channel Migration Area (area Acres 0.0 60 122 124 43 between existing levee and setback levee) Reconnected Floodplain Area within Acres 0 45 74 70 23 100-year Inundation Area Reconnected Floodplain Area within Acres 0 34 49 46 20 2-year Inundation Area Carbon River Potential Active Channel Width at RM 3.8 Pinch Point Feet 260 540 540 850 380 (assuming right bank side remains fixed) HABITAT Side channel created Lineal Feet 0.0 3500 3500 3600 600 Existing Metric Units Alt. 1 Alt. 2 Alt. 3 Alt. 4 Conditions 26 multi-log jams and 583 individual Wood added to active Number key logs 700 750 750 300 channel pieces (Per analysis of 2018 aerial photograph) Wood added to Number key Not 200 250 250 100 floodplain pieces measured Wetlands Impacted by Acres 0.0 7 5 5 4 Levee Footprint Floodplain Riparian and Wetland Acres 0.0 45 61 60 25 Establishment or Enhancement VOIGHTS CREEK Voights Creek Reach Length in Same as Reconnected Miles 0.00 0.16 0.57 0.39 existing Floodplain (Setback condition Levee to Existing Levee) Voights Creek Length Same as SR162 to Setback Miles 0.57 0.41 0.0 0.18 existing Levee condition Crossing at New Crossing New Crossing existing at Setback at Setback levee No new Retains Fish passable Levee Levee -- appears to crossing existing crossings replaces replaces meet WDFW required. -

A Geomorphic Classification System
A Geomorphic Classification System U.S.D.A. Forest Service Geomorphology Working Group Haskins, Donald M.1, Correll, Cynthia S.2, Foster, Richard A.3, Chatoian, John M.4, Fincher, James M.5, Strenger, Steven 6, Keys, James E. Jr.7, Maxwell, James R.8 and King, Thomas 9 February 1998 Version 1.4 1 Forest Geologist, Shasta-Trinity National Forests, Pacific Southwest Region, Redding, CA; 2 Soil Scientist, Range Staff, Washington Office, Prineville, OR; 3 Area Soil Scientist, Chatham Area, Tongass National Forest, Alaska Region, Sitka, AK; 4 Regional Geologist, Pacific Southwest Region, San Francisco, CA; 5 Integrated Resource Inventory Program Manager, Alaska Region, Juneau, AK; 6 Supervisory Soil Scientist, Southwest Region, Albuquerque, NM; 7 Interagency Liaison for Washington Office ECOMAP Group, Southern Region, Atlanta, GA; 8 Water Program Leader, Rocky Mountain Region, Golden, CO; and 9 Geology Program Manager, Washington Office, Washington, DC. A Geomorphic Classification System 1 Table of Contents Abstract .......................................................................................................................................... 5 I. INTRODUCTION................................................................................................................. 6 History of Classification Efforts in the Forest Service ............................................................... 6 History of Development .............................................................................................................. 7 Goals -

The Favorability of Florida's Geology to Sinkhole
Appendix H: Sinkhole Report 2018 State Hazard Mitigation Plan _______________________________________________________________________________________ APPENDIX H: Sinkhole Report _______________________________________________________________________________________ Florida Division of Emergency Management THE FAVORABILITY OF FLORIDA’S GEOLOGY TO SINKHOLE FORMATION Prepared For: The Florida Division of Emergency Management, Mitigation Section Florida Department of Environmental Protection, Florida Geological Survey 3000 Commonwealth Boulevard, Suite 1, Tallahassee, Florida 32303 June 2017 Table of Contents EXECUTIVE SUMMARY ............................................................................................................ 4 INTRODUCTION .......................................................................................................................... 4 Background ................................................................................................................................. 5 Subsidence Incident Report Database ..................................................................................... 6 Purpose and Scope ...................................................................................................................... 7 Sinkhole Development ................................................................................................................ 7 Subsidence Sinkhole Formation .............................................................................................. 8 Collapse Sinkhole -

Restoring a Degraded Gulf of Mexico
April 2012 | Visit: www.NWF.org/oilspill Introduction Three years after the Deepwater Horizon disaster, this report gives a snapshot view of six wildlife species that depend on a healthy Gulf and the coastal wetlands that are critical to the Gulf’s food web. In August 2011, scientists did a comprehensive health examination of a 16-year-old male bottlenose dolphin. This dolphin—dubbed Y12 for research purposes—was caught near Grand Isle, a Louisiana barrier island that was oiled during the Gulf oil disaster.i Like many of the 31 other dolphins examined in the study, Y12 was found to be severely ill—underweight, anemic and with signs of liver and lung disease. The dolphins’ symptoms were consistent with those seen in other mammals exposed to oil; researchers feared many of the dolphins studied were so ill they would not survive.ii Seven months later, Y12’s emaciated carcass washed up on the beach at Grand Isle.iii Ecosystem-wide effects of the oil are suggested by the poor health of dolphins, which are at the top of the Gulf’s food chain. The same may be true of sea turtles, which also continue to die in alarmingly high numbers. More than 650 dolphins have been found stranded in the oil spill area since the Gulf oil disaster began. This is more than four times the historical average. Research shows that polycyclic aromatic hydrocarbon (PAH) components of oil from the Macondo well were found in plankton even after the well was capped.iv, v PAHs can have carcinogenic, physiological and genetic effects. -

Cynthia Winings Gallery
ROAD WArrIOR Swept Away We interrupt this magazine for OM C O. C a live feed directly from the Secretary of Defense’s house. HELL; POLITI C IT BY COLIN W. SARGENT M uring the Reagan presidency, everyone on Earth knew that Secretary of De- fense Caspar Weinberger (1917-2006) MILY CARTER E D kept a getaway place up in Maine. But it was more something imagined than ac- tually seen. Far fewer knew where it was, or how luxuriant it was, or that it was once owned by Mary King Auchincloss. Listed for sale this sum- HE KOWLES COMPANY; T mer for $2.15M, “Windswept,” built in 1910, en- joys a classic view of Somes Sound on Mount FROM TOP: Desert Island. Positioned to advantage at 584 SUMMERGUIDE 2 0 1 6 1 4 7 A Chat With Caspar W. Weinberger, Jr. Caspar Willard Weinberger, Jr, known as “Cap,” is the son of Caspar Weinberger, prominent statesman and, most notably, U.S. Secretary of Defense under the Rea- gan administration from 1981 to 1987. Cap’s mother, Jane Weinberger, was an author and publisher from Milford, Maine. Born in 1947, Cap studied at Harvard–his father’s alma mater–and earned a B.A. in Modern British History in 1968. He lived in San Francisco for many years, working as an award-winning producer, director, and writer of documentary films for an NBC affiliate. Cap settled in Mount Desert in 1997. Today he is a writer and lecturer, as well as running Windswept House Publish- ers, started by his mother in 1984.