(Thismiaceae) from Ulu Temburong, Brunei Darussalam
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Belait District
BELAIT DISTRICT His Majesty Sultan Haji Hassanal Bolkiah Mu’izzaddin Waddaulah ibni Al-Marhum Sultan Haji Omar ‘Ali Saifuddien Sa’adul Khairi Waddien Sultan and Yang Di-Pertuan of Brunei Darussalam ..................................................................................... Kebawah Duli Yang Maha Mulia Paduka Seri Baginda Sultan Haji Hassanal Bolkiah Mu’izzaddin Waddaulah ibni Al-Marhum Sultan Haji Omar ‘Ali Saifuddien Sa’adul Khairi Waddien Sultan dan Yang Di-Pertuan Negara Brunei Darussalam BELAIT DISTRICT Published by English News Division Information Department Prime Minister’s Office Brunei Darussalam BB3510 The contents, generally, are based on information available in Brunei Darussalam Newsletter and Brunei Today First Edition 1988 Second Edition 2011 Editoriol Advisory Board/Sidang Redaksi Dr. Haji Muhammad Hadi bin Muhammad Melayong (hadi.melayong@ information.gov.bn) Hajah Noorashidah binti Haji Aliomar ([email protected]) Editor/Penyunting Sastra Sarini Haji Julaini ([email protected]) Sub Editor/Penolong Penyunting Hajah Noorhijrah Haji Idris (noorhijrah.idris @information.gov.bn) Text & Translation/Teks & Terjemahan Hajah Apsah Haji Sahdan ([email protected]) Layout/Reka Letak Hajah Apsah Haji Sahdan Proof reader/Penyemak Hajah Norpisah Md. Salleh ([email protected]) Map of Brunei/Peta Brunei Haji Roslan bin Haji Md. Daud ([email protected]) Photos/Foto Photography & Audio Visual Division of Information Department / Bahagian Fotografi -

Preliminary Report of BPP 2011
! ! ! Kerajaan!Kebawah!Duli!Yang!Maha!Mulia!Paduka!Seri!Baginda!Sultan!dan!Yang!Di8Pertuan! Negara! Brunei! Darussalam! melalui! Jabatan! Perancangan! dan! Kemajuan! Ekonomi! (JPKE),! Jabatan!Perdana!Menteri,!telah!mengendalikan!Banci!Penduduk!dan!Perumahan!(BPP)!pada! tahun! 2011.! BPP! 2011! merupakan! banci! kelima! seumpamanya! dikendalikan! di! negara! ini.! Banci!terdahulu!telah!dijalankan!pada!tahun!1971,!1981,!1991!dan!2001.! ! Laporan! Awal! Banci! Penduduk! dan! Perumahan! 2011! ini! merupakan! penerbitan! pertama! dalam! siri! laporan8laporan! banci! yang! akan! dikeluarkan! secara! berperingkat8peringkat.! Laporan! ini! memberikan! data! awal! mengenai! jumlah! penduduk,! isi! rumah! dan! tempat! kediaman!serta!taburan!dan!pertumbuhan!mengikut!daerah.!! ! Saya! berharap! penerbitan! ini! dan! laporan8laporan! seterusnya! akan! dapat! memenuhi! keperluan! pelbagai! pengguna! di! negara! ini! bagi! maksud! perancangan,! penyelidikan,! penyediaan!dasar!dan!sebagai!bahan!rujukan!awam.! ! Saya! sukacita! merakamkan! setinggi8tinggi! penghargaan! dan! terima! kasih! kepada! Penerusi! dan! ahli8ahli! Komiti! Penyelarasan! Kebangsaan! BPP! 2011,! kementerian8kementerian,! jabatan8jabatan! dan! sektor! swasta! yang! telah! memberikan! bantuan! dan! kerjasama! yang! diperlukan! kepada! Jabatan! ini! semasa! banci! dijalankan.! Seterusnya! saya! juga! sukacita! mengucapkan! terima! kasih! kepada! rakyat! dan! penduduk! di! negara! ini! di! atas! kerjasama! dalam! memberikan! maklumat! yang! dikehendaki! kepada! pegawai8pegawai! banci! -

Brunei Darussalam
Published by: Ministry of Finance and Economy Commonwealth Drive, Bandar Seri Begawan BB3910 Brunei Darussalam www.mofe.gov.bn In collaboration with: Authority for Info-communications Technology Industry of Brunei Darussalam Autoriti Monetari Brunei Darussalam Brunei Intellectual Property Office Employees Trust Fund Ministry of Culture, Youth and Sports Ministry of Defence Ministry of Development Ministry of Education Ministry of Energy Ministry of Foreign Affairs Ministry of Health Ministry of Home Affairs Ministry of Primary Resources and Tourism Ministry of Religious Affairs Ministry of Transport and Infocommunications Prime Minister’s Office Wawasan Brunei 2035 Standing Secretariat Bank Islam Brunei Darussalam Biodiversity and Natural History Society Brunei Council on Social Welfare Brunei Darussalam AIDS Council Brunei Darussalam National Association of the Blind Brunei Darussalam Youth Council Brunei Malay Chamber of Commerce and Industry Chinese Chamber of Commerce in Brunei Darussalam Council for the Welfare of Persons with Different Abilities Down Syndrome Association Green Brunei Hand4Hand Brunei La Vida Sdn Bhd MyActionsforSDGs National Chamber of Commerce and Industry Brunei Darussalam Pusat Ehsan Al-Ameerah Al-Hajjah Maryam Society for Community Outreach and Training Society for the Management of Autism Related Issues in Training, Education and Resources Sultan Haji Hassanal Bolkiah Foundation Universiti Brunei Darussalam WeCare | KamiPrihatin © Ministry of Finance and Economy, Government of Brunei Darussalam 2020. All rights -
![[PREVIEW] Brunei Temburong Link](https://docslib.b-cdn.net/cover/4291/preview-brunei-temburong-link-2244291.webp)
[PREVIEW] Brunei Temburong Link
IABSE Conference – Engineering the Developing World April 25-27 2018, Kuala Lumpur, Malaysia Brunei Temburong Link Naeem Hussain, Sammy Yip, Murphy To Ove Arup & Partners, Hong Kong Chin Kok Kong, Ding Lee Sing, Yat Kheong Cheng Ove Arup Dan Rakan-Rakan, Brunei Contact: [email protected] Abstract Temburong district has been physically isolated from the rest of Brunei which has hindered the economic growth of the district. A sea-crossing link is being constructed to connect the district to Brunei-Muara. A feasibility study was carried out between 2010 and 2012 to establish the alignment and preliminary design of the link, followed by detail design and construction. This paper describes aspects of the feasibility study, detail design and construction Keywords: Alignment, marine viaduct, swamp viaduct, cable-stay bridges, pre-cast construction. Bay and then up-river to Bangar. Neither of these 1 Introduction alternatives offer 24 hours a day accessibility. Brunei is located on the north coast of the island of Borneo in Southeast Asia. Apart from its coastline with the South China Sea, it is completely surrounded by the state of Sarawak, Malaysia; and is separated into two parts by the Sarawak district of Limbang, Figure 1. This physical separation of Temburong district from the majority of Brunei has significantly affects the economic development of the district and Brunei as a whole. Despite comprising 23% of the total land area of Brunei, Temburong only contains 2% of the population. To travel to Temburong by land requires passing into and out of Sarawak through a long circuitous route typically involving significant waiting times at the borders which can result in a Figure 1. -

Brunei Darussalam
Country profile – Brunei Darussalam Version 2011 Recommended citation: FAO. 2011. AQUASTAT Country Profile – Brunei Darussalam. Food and Agriculture Organization of the United Nations (FAO). Rome, Italy The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of specific companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned. The views expressed in this information product are those of the author(s) and do not necessarily reflect the views or policies of FAO. FAO encourages the use, reproduction and dissemination of material in this information product. Except where otherwise indicated, material may be copied, downloaded and printed for private study, research and teaching purposes, or for use in non-commercial products or services, provided that appropriate acknowledgement of FAO as the source and copyright holder is given and that FAO’s endorsement of users’ views, products or services is not implied in any way. All requests for translation and adaptation rights, and for resale and other commercial use rights should be made via www.fao.org/contact-us/licencerequest or addressed to [email protected]. FAO information products are available on the FAO website (www.fao.org/ publications) and can be purchased through [email protected]. -

Brunei: Toponymic Factfile (August 2018)
TOPONYMIC FACT FILE Brunei Country name Brunei State title Brunei Darussalam Official language(s) Malay (msa)1 Brunei / Brunei Darussalam2 Country name in official language(s) بروني دارالسﻻم / بروني Negara Brunei Darussalam3 State title in official language(s) نڬارا بروني بروني /دارالسﻻم Malay is written in Rumi (Roman-script) or Jawi Script (modified Arabic-script)4 Romanization System Not required ISO-3166 code (alpha-2/alpha-3) BN/BRN Capital(s) Bandar Seri Begawan5 Introduction The state of Brunei is situated on the north-western coast of the island of Borneo on the South China Sea. It is surrounded on all other sides by the Malaysian State of Sarawak which divides Brunei into two unconnected parts: the eastern part of which is the rural Temburong district, and the western part which consists of Brunei Muara, Tutong and Belait districts. Brunei acquired full independence from the United Kingdom in 1984. Geographical names policy Recommended geographical names are those in Roman-script as found on official Brunei-produced sources, where possible. The National Committee of Geographical Names Jawatankuasa Kebangsaan Nama Geografi (JKNG), deals with the standardisation of place names in Brunei. Official maps are produced by the Brunei Survey Department. The Brunei Survey Department’s Geoportal6 can be used to access geospatial information (including geographical names) online. 1 It is called Bahasa Melayu ("Malay language") in Malay. Language codes given in this Factfile are ISO-639 codes. 2 The names Brunei and Brunei Darussalam are used interchangeably in Brunei. 3 No state title as such occurs in the Brunei Constitution of 1959, but the name Negara Brunei Darussalam (meaning Nation of Brunei, the Abode of Peace) was proclaimed as the official name of the state in June 1983. -

Geo-Data: the World Geographical Encyclopedia
Geodata.book Page iv Tuesday, October 15, 2002 8:25 AM GEO-DATA: THE WORLD GEOGRAPHICAL ENCYCLOPEDIA Project Editor Imaging and Multimedia Manufacturing John F. McCoy Randy Bassett, Christine O'Bryan, Barbara J. Nekita McKee Yarrow Editorial Mary Rose Bonk, Pamela A. Dear, Rachel J. Project Design Kain, Lynn U. Koch, Michael D. Lesniak, Nancy Cindy Baldwin, Tracey Rowens Matuszak, Michael T. Reade © 2002 by Gale. Gale is an imprint of The Gale For permission to use material from this prod- Since this page cannot legibly accommodate Group, Inc., a division of Thomson Learning, uct, submit your request via Web at http:// all copyright notices, the acknowledgements Inc. www.gale-edit.com/permissions, or you may constitute an extension of this copyright download our Permissions Request form and notice. Gale and Design™ and Thomson Learning™ submit your request by fax or mail to: are trademarks used herein under license. While every effort has been made to ensure Permissions Department the reliability of the information presented in For more information contact The Gale Group, Inc. this publication, The Gale Group, Inc. does The Gale Group, Inc. 27500 Drake Rd. not guarantee the accuracy of the data con- 27500 Drake Rd. Farmington Hills, MI 48331–3535 tained herein. The Gale Group, Inc. accepts no Farmington Hills, MI 48331–3535 Permissions Hotline: payment for listing; and inclusion in the pub- Or you can visit our Internet site at 248–699–8006 or 800–877–4253; ext. 8006 lication of any organization, agency, institu- http://www.gale.com Fax: 248–699–8074 or 800–762–4058 tion, publication, service, or individual does not imply endorsement of the editors or pub- ALL RIGHTS RESERVED Cover photographs reproduced by permission No part of this work covered by the copyright lisher. -

Agriculture and Agrifood Statistic 2020
Brunei Darussalam Agriculture & Agrifood Statistics in Brief 2020 1 First Publication. 2021. JP/BPPA/01/2021 PREFACE The year 2020 has been a challenging year for Brunei The statistical data this year has shown positive growth, the Darussalam and globally due to the outbreak of Coronavirus gross output of the agriculture and agrifood sector for 2020 has with many sectors and industries affected, and Agriculture increase 9.94% valued at B$470.86 million, of which B$267.17 and Agrifood Sector is no exception. Alhamdulillah, our million comes from the livestock industry; B$53.46 million country managed to contain this challenge as a result of from the crop industry; and B$150.23 million from the agrifood concerted efforts by relevant parties, including entrepreneurs processing industry. It is hoped that continuous growth will be in the primary resources sector in terms of increasing food able to support the country’s goals and aspirations to increase production, as well as the involvement and cooperation of all the level of self-sufficiency and food security in the country. with the ‘whole of nation’ approach. The Department would like to convey its sincere appreciation to Despite the challenges faced in 2020, the Department of all relevant officers and staff of the Department of Agriculture Agriculture and Agrifood has successfully conducted new and Agrifood (DOAA) for their continuous assistance and programs and improvement efforts to increase production in support in providing the relevant data. The Department will the agricultural sector, such as the introduction of the Pasar continue to improve this publication from time to time in line Kitani Program which is to help our local farmers market their with the increasing demand and current statistical development. -

Ministry of Finance and Economy Brunei Darussalam
MINISTRY OF FINANCE AND ECONOMY BRUNEI DARUSSALAM PRESS RELEASE IMPLEMENTATION OF EXIT AND ENTRY CHARGES ORDER, 2020 THURSDAY, 17 ZULKAEDAH 1441 CORRESPONDING TO 09 JULY 2020 1. By command of His Majesty Sultan Haji Hassanal Bolkiah Mu`izzaddin Waddaulah ibni Al-Marhum Sultan Haji Omar `Ali Saifuddien Sa`adul Khairi Waddien, Sultan and Yang Di-Pertuan of Brunei Darussalam, the Ministry of Finance and Economy hereby announces that His Majesty has consented to the implementation of the Exit and Entry Charges Order, 2020 which will be enforced on 1st August 2020. 2. This Order gives the Controller of Customs and Excise, Royal Customs and Excise Department the power to charge any person who intends to leave or enter Brunei Darussalam through the land control posts (checkpoints). 3. The main objective for the implementation of these charges, named ‘Caj Perkhidmatan’ (CaP) is to standardize the imposition of payments or charges that are imposed to the public upon either leaving or entering Brunei Darussalam via air and sea. With the enforcement of this Order, toll charges at Rasau Bridge in the Belait District will be eliminated on the date mentioned earlier. 1 4. This charge will be imposed to all citizens including Brunei Darussalam citizens who intend to leave or enter Brunei Darussalam through the land control posts (checkpoints) either via vehicle or as a pedestrian with a charge rate of BND3.00 per person for each one-way trip. For example, a return trip to Miri, Sarawak will cost BND6.00 per person. 5. The charge will not be applicable to persons or class of persons as below: 5.1 Occupants of the following vehicles: ● A vehicle which belongs to the Government of His Majesty the Sultan and Yang Di-Pertuan of Brunei Darussalam or a government of another country when it is being used for official purpose; ● A vehicle which belongs to the military forces of Brunei Darussalam; ● A vehicle when it is being used for firefighting, ambulance or police purposes; and ● A vehicle when it is being used for the purpose of a funeral. -

Chapter 2. Analysis and Assessment of Current Conditions
Chapter 2 Analysis and Assessment of Current Conditions 2.1 Brunei Vision 2035 (Wawasan Brunei 35) Wawasan Brunei 2035 is the vision of the nation towards 2035 announced by the Government of Brunei Darussalam in January 2008. It aims to turn Brunei into a nation widely recognised for its quality of life amongst the top 10 nations in the world, its well-educated and highly-skilled people, and a dynamic and sustainable economy–without relying heavily on oil and natural gas. To realise this vision, the long-term development strategy and outline for policies served as the guideline for the next 10 years’ development. Projects are identified based on the five-year national development plan. Figure 2.1: Sites of Main Development Project Lugu: Meragang: Lambak Kanan Easet: New technology Logistic, warehouse Food, medicine Bukit Panggal: Agrotech Park: Pulau Muara Besar: Renewable energy Agriculture, Food Petroleum processing Petrochemistry, Oil gas Teliai: Satellite・gateway telecommunication Bandar Seri Begawan: Financial center Sg .Liang Industrial Park: Petro cmistry Temburong Seria: Petroleum & Natural Tasek Merimbun: Ulu-Ulu: Gas Ecotourism Ecotourism Source: Study team. In February 2017, the government announced the next phase of economic policies, reflecting progress towards Wawasan Brunei 2035. These policies will (i) stimulate other industries to overcome the dependence on oil and natural gas, (ii) implement measures to support domestic entrepreneurs, (iii) implement incentives for small and medium-sized enterprises, and (iv) make effective use of them when ASEAN Economic Community is established. 5 2.2 Heart of Borneo 2.2.1 Natural Environment of Borneo Island The island of Borneo has one of the world’s most biologically diverse tropical rainforests, lying just below the equator. -
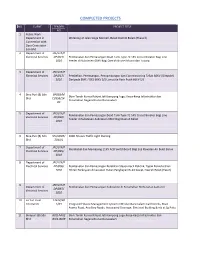
Completed Projects
COMPLETED PROJECTS NO CLIENT TENDER PROJECT TITLE NO 1 Public Work Department in Widening of Jalan Singa Menteri, Belait District Belait (Phase ll) Connection with Dara Contractor Sdn Bhd 2 Department of JPE/LTK/P Electrical Services DP/023/ Pembekalan dan Pemasangan Dead Tank Type 72.5KV Circuit Breaker Bagi Line 2010 Feeder di Substesen 66KV Bagi Daerah Brunei Muara dan Tutong 3 Department of JPE/LTK/P Electrical Services DP/017/ Pembelian, Pemasangan, Penyambungan dan Commissioning Talian 66KV (Slimpole) 2010 Daripada BMC / DES 66KV S/S Lumut Ke Pasir Putih 66KV S/S 4 Bina Puri (B) Sdn BPBSB/M Skim Tanah Kurnia Rakyat Jati Kampung Lugu: Kerja-Kerja Infastruktur dan Bhd CI/SSB/14- Perumahan Negara Brunei Darussalam 03 5 Department of JPE/LTK/P Pembekalan dan Pemasangan Dead Tank Type 72.5KV Circuit Breaker Bagi Line Electrical Services DP/030/ Feeder di Substesen-Substesen 66KV Bagi Daerah Belait 2010 6 Bina Puri (B) Sdn SIV/Q005/ 2000 Houses Traffic Light Ducting Bhd 2011/L 7 Department of JPE/LTK/P Membekal dan Memasang 11KV ACB Switchboard Bagi Loji Rawatan Air Bukit Barun Electrical Services DP/095/ 2010 8 Department of JPE/LTK/P Electrical Services DP/096/ Pembekalan dan Pemasangan Peralatan Stesen Kecil Elektrik, Tapak Perindustrian 2010 Hiliran Perkayuan di kawasan Hutan Pengkajian Bukit Sawat, Daerah Belait (Fasa1) 9 JPE/LTK/P Department of Pembekalan dan Pemasangan Substation di Perumahan Perhutanan Lamunin DP/097/ Electrical Services 2010 10 Fu Yun Yuan 13612/20 Contractor 1/03 Integrated Waste Management System in Brunei -

Brunei: the ASEAN Connection Donald E
University of South Carolina Scholar Commons Faculty Publications Political Science, Department of 6-1983 Brunei: The ASEAN Connection Donald E. Weatherbee University of South Carolina - Columbia, [email protected] Follow this and additional works at: https://scholarcommons.sc.edu/poli_facpub Part of the Political Science Commons Publication Info Published in Asian Survey, Volume 23, Issue 6, 1983, pages 723-735. http://www.ucpressjournals.com/journal.asp?j=as © 1983 by University of California Press This Article is brought to you by the Political Science, Department of at Scholar Commons. It has been accepted for inclusion in Faculty Publications by an authorized administrator of Scholar Commons. For more information, please contact [email protected]. BRUNEI TheASEAN Connection Donald E. Weatherbee Although no formal mechanism for accession has yet been announced, it is expected by Brunei and its future partners that the Sultanate will become the sixth member of the Association of Southeast Asian Nations (ASEAN) in 1984, after it obtains full sovereignty over its external affairs.'The characteristicASEAN decision-making process of for- mal and informalconsultations (musyawarah)leading to consensus (mufakat) is in full train. Brunei'scandidature was announced at the June 1981 annual ASEAN Foreign Ministers meeting in Manila. Brunei was invited as an "observer"and was representedby the Sultan's brother, Prince Mohammad Bolkiah, who is the PerdanaWazir and probably will be Brunei's first For- eign Minister. In his address to the conference, Prince Mohammad stated that it was the hope of Brunei that the question of its membership in ASEAN could be favorably resolved once independence was achieved.2 If the statements of ASEAN leaders to date are any guide, there would appear to be little doubt that when Brunei makes its application, it will be accepted.