Final Conference Programme Now Available As
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Crapemyrtle Bark Scale Acanthococcus (=Eriococcus) Lagerstroemiae
Crapemyrtle Bark Scale Acanthococcus (=Eriococcus) lagerstroemiae Jim Robbins, Univ. of Ark. CES, Bugwood.org, #5521505 The crapemyrtle bark scale (Acanthococcus (=Eriococcus) lagerstroemiae) is in the family Eriococcidae (or Acanthococcidae, as the taxonomy of this family is still being debated). This group is in the superfamily Coccoidea (scale insects) and the order Hemiptera (true bugs). The primary host in North America, crapemyrtle, Lagerstroemia spp., are deciduous flowering trees popular in ornamental landscapes. They are top-sellers in the nursery trade, with the annual wholesale value estimated to be $66 million in 2014. Based on urban tree inventories of several major cities in the southeastern U.S., crapemyrtle are among the most common landscape trees planted in this region. Resources - Cai, X., H. Dou, M. Gu, M. Merchant, and E. Vafaie. 2015. Update on crapymyrtle bark scale. Proceedings of the 2015 Annual Meeting of the International Plant Propagators’ Society. 1140: 415–418. - Wang, Z., Y. Chen, M. Gu, E. Vafaie, M. Merchant, and R. Diaz. 2016. Crapemyrtle bark scale: A new threat for crapemyrtles, a popular landscape plant in the U.S. Insects. 7: 33–34. 1 Crapemyrtle Bark Scale Close-up of a crapemyrtle bark scale • In the group of insects infestation, showing the fuzzy, felt-like known as felt scales covering of the adult female scales • Native to Asia • First report in the US – 2004 in Richardson, Texas (Dallas County) • Formerly known as Eriococcus lagerstroemiae Kuwana Gary Brooks, Bayer CropScience, Bugwood.org #5523058 The crapemyrtle bark scale (abbreviated as CMBS) belongs to a group of scale insects known as felt scales or bark scales. -

Ist Β-Glucosidase Ein Schlüsselenzym Für Die
Ist β-Glucosidase ein Schlüsselenzym für die Anpassung an Iridoidglycoside? Ein Vergleich zwischen herbivoren Generalisten (Arctiidae, Lepidoptera) und einem Iridoidglycosidspezialist (Nymphalidae, Lepidoptera) Dissertation zur Erlangung der Doktorgrades der Naturwissenschaften (Dr. rer. nat.) im Department Biologie der Fakultät für Mathematik, Informatik und Naturwissenschaften an der Universität Hamburg vorgelegt von Helga Carola Pankoke aus Freiburg im Breisgau Hamburg 2010 Erstgutachterin: Professor Dr. Susanne Dobler Zweitgutachter: Professor Dr . Reinhard Lieberei Tag der Disputation: 9. April 2010 Bestätigung zur sprachlichen Korrektheit der in englischer Sprache abgefassten Dissertation Die sprachliche Korrektheit der in englischer Sprache abgefassten Dissertation von Frau Helga Carola Pankoke mit wurde von der Unterzeichnenden geprüft und bestätigt. (engl . „The grammatical and linguistical correctness of the dissertation, which has been written in English by Ms. Helga Carola Pankoke, has been proved and confirmed.“) (M. Deane Bowers) Museum and Department of Ecology and Evolutionary Biology, UCB 334 University of Colorado, Boulder Colorado 80309, USA Inhaltsverzeichnis Zusammenfassung i 1 Allgemeine Einleitung 01 2 Influence of iridoid glycoside containing host plants on midgut β-glucosidase activity in the yellow woolly bear, Spilosoma virginica (Arctiidae) 06 2.1 Summary 06 2.2 Introduction 07 2.3 Materials and Methods 10 2.4 Results 14 2.5 Discussion 19 3 The interplay between toxin-releasing β-glucosidase and plant iridoid -

Disease Resistant Ornamental Trees
HOME GROUNDS FACT SHEET Horticulture Center Cornell University Demonstration & Community Gardens at East Meadow Farm Cooperative Extension 832 Merrick Avenue East Meadow, NY 11554 Nassau County Phone: 516-565-5265 Disease-Resistant Ornamental Trees and Those Relatively Free of Disease Some trees show resistance to certain of the more Flowering crabapples serious tree diseases. Others are relatively free from * Malus ‘Adams’ o major disease problems under growing conditions found Malus baccata var. himalaica in the Northeast. Many of these latter types are used Malus ‘Cardinal’ All unmarked crab- only occasionally in shade and ornamental plantings, a * Malus ‘Centurion’ o apples are resistant to Malus ‘David’ cedar apple rust, fire fact that may account for the general absence of dis- Malus ‘Donald Wyman’ blight, powdery mildew and scab. eases. However, with the increased limitations on the * Malus floribunda o use of effective fungicides for the control of tree dis- Malus ‘Henry Kohankie’ o Crabapples suscep- tible to one or more eases, it would be advisable to check the lists below Malus ‘Inglis’ of the mentioned dis- and substitute one of these trees for a tree that is Malus ‘Professor Sprenger’ eases, but still worth susceptible to diseases. * Malus ‘Profusion’ planting. Malus ‘Robinson’ * Indicates varieties * Malus sargentii o commonly available on Long Island. Resistant to Dutch elm disease: * Malus ‘Sugar Tyme’ o Ulmus parvifolia (Chinese elm) Ulmus glabra 'camperdownii' (Scotch elm) Junipers resistant to twig blight caused by Resistant to chestnut blight, but Phomopsis juniperovara: susceptible to twig blight and canker: Juniperus chinensis Castanea mollissima (Chinese chestnut) ‘Foemina’ ++ Castanea crenata (Japanese chestnut) ‘Iowa’ ‘Keteleeri’ ++ Resistant to witch’s broom: * ‘Robusta green' Celtis sinensis (Chinese hackberry) * var. -

Taxonomic Overview of Ligustrum (Oleaceae) Naturalizaed in North America North of Mexico
Phytologia (December 2009) 91(3) 467 TAXONOMIC OVERVIEW OF LIGUSTRUM (OLEACEAE) NATURALIZAED IN NORTH AMERICA NORTH OF MEXICO Guy L. Nesom 2925 Hartwood Drive Fort Worth, TX 76109, USA www.guynesom.com ABSTRACT A key, morphological descriptions, and basic synonymy are provided for the eight species of Ligustrum known to be naturalized in North America north of Mexico: L. japonicum, L. lucidum, L. obtusifolium (including L. amurense), L. ovalifolium, L. quihoui, L. sinense, L. tschonoskii, and L. vulgare. Identifications have been inconsistent particularly between L. sinense and L. vulgare and between L. japonicum and L. lucidum. The occurrence of L. quihoui outside of cultivation in Arkansas, Mississippi, and Oklahoma is documented. Phytologia 91(3): 467-482 (December, 2009). KEY WORDS: Ligustrum, Oleaceae, North America, naturalized, taxonomy The lustrous, mostly evergreen leaves and masses of white, fragrant flowers make privets popular for landscaping and hedges. Many of the species, however, have become naturalized in the USA and Canada and already have proved to be destructive colonizers, especially in the Southeast. Among the naturalized species, European privet (Ligustrum vulgare) is native to Europe and northern Africa; all the rest are native to Asia, mainly China, Japan, and Korea. Many new species and varieties of Ligustrum have been described since overviews of Koehne (1904), Lingelsheim (1920), and Mansfield (1924). The genus in eastern Asia has recently been studied by Chang & Miao (1986), and Qin (2009) has provided a taxonomic overview of the whole genus that recognizes 37 species - divided into five sections based primarily on fruit and seed morphology. In Qin’s arrangement, among the North American species, sect. -

Non-Native Invasive Plants of the City of Alexandria, Virginia
March 1, 2019 Non-Native Invasive Plants of the City of Alexandria, Virginia Non-native invasive plants have increasingly become a major threat to natural areas, parks, forests, and wetlands by displacing native species and wildlife and significantly degrading habitats. Today, they are considered the greatest threat to natural areas and global biodiversity, second only to habitat loss resulting from development and urbanization (Vitousek et al. 1996, Pimentel et al. 2005). The Virginia Department of Conservation and Recreation has identified 90 non-native invasive plants that threaten natural areas and lands in Virginia (Heffernan et al. 2014) and Swearingen et al. (2010) include 80 plants from a list of nearly 280 non-native invasive plant species documented within the mid- Atlantic region. Largely overlapping with these and other regional lists are 116 species that were documented in the City of Alexandria, Virginia during vegetation surveys and natural resource assessments by the City of Alexandria Dept. of Recreation, Parks, and Cultural Activities (RPCA), Natural Lands Management Section. This list is not regulatory but serves as an educational reference informing those with concerns about non-native invasive plants in the City of Alexandria and vicinity, including taking action to prevent the further spread of these species by not planting them. Exotic species are those that are not native to a particular place or habitat as a result of human intervention. A non-native invasive plant is here defined as one that exhibits some degree of invasiveness, whether dominant and widespread in a particular habitat or landscape or much less common but long-lived and extremely persistent in places where it occurs. -

Identification and Control of Invasive Privets (Ligustrum Spp.) in the Middle Southern United States
Invasive Plant Science and Management 2010 3:482–488 Notes and Commentary Identification and Control of Invasive Privets (Ligustrum spp.) in the Middle Southern United States Victor Maddox, John Byrd, Jr., and Brett Serviss* The identification of privet in the middle southern United States can be difficult. Because most introduced species of privet can be invasive, and recent mapping projects seek location and species population data, proper identification is important. Without proper identification of privet species, data on species distributions and other pertinent information regarding invasiveness could lead to improper conclusions. Currently, information on privet identification is scattered throughout a number of reference materials. The purpose of this publication is to assist with the proper identification of escaped privet species, and suggest management options. Nomenclature: Fosamine ammonium; glyphosate; hexazinone; imazapyr; metsulfuron; triclopyr; 2,4-D; 2,4-DP; Amur privet, Ligustrum obtusifolium Sieb. & Zucc. var. suave (Kitagawa) Kitagawa (Syn. L. amurense Carrie`re); border privet, Ligustrum obtusifolium Sieb. & Zucc. var. obtusifolium; California privet, Ligustrum ovalifolium Hassk.; Chinese privet, Ligustrum sinense Lour.; common privet, Ligustrum vulgare L.; glossy privet, Ligustrum lucidum Ait.; Japanese privet, Ligustrum japonicum Thunb.; waxyleaf privet, Ligustrum quihoui Carrie`re. Key words: Invasive species, management. Since the 1700s, at least nine species of privets have been Thunb.) is native to Korea and Japan. The most common introduced into the United States; it is probable that all species in the southern portion of the middle southern were introduced as ornamentals. They have been very region is Chinese privet, although Amu, border, California, successful as ornamentals and continue to be marketed for common or European, glossy, Japanese, and waxyleaf or such purposes. -

Ligustrum Sinense LOUR.)
CAUSES AND CONSEQUENCES OF CHINESE PRIVET (Ligustrum sinense LOUR.) INVASION IN HYDROLOGICALLY ALTERED FORESTED WETLANDS Meghan Foard A thesis presented to the faculty of Arkansas State University in partial fulfillment of the requirements of the degree of MASTER OF SCIENCE IN ENVIRONMENTAL SCIENCE Arkansas State University August 2014 Approved by Dr. Travis D. Marsico, Thesis Advisor Dr. Jennifer Bouldin Committee Member Dr. Richard Grippo, Committee Member Dr. Esra Ozdenerol, Committee Member Dr. Paul Sikkel, Committee Member UMI Number: 1563273 All rights reserved INFORMATION TO ALL USERS The quality of this reproduction is dependent upon the quality of the copy submitted. In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed, a note will indicate the deletion. UMI 1563273 Published by ProQuest LLC (2014). Copyright in the Dissertation held by the Author. Microform Edition © ProQuest LLC. All rights reserved. This work is protected against unauthorized copying under Title 17, United States Code ProQuest LLC. 789 East Eisenhower Parkway P.O. Box 1346 Ann Arbor, MI 48106 - 1346 © 2014 Meghan Foard ALL RIGHTS RESERVED ii ABSTRACT Meghan Foard CAUSES AND CONSEQUENCES OF CHINESE PRIVET (Ligustrum sinense LOUR.) INVASION IN HYDROLOGICALLY ALTERED FORESTED WETLANDS What drives invasive species success? My research consists of four studies aiming to answer this question for Ligustrum sinense. The four projects are: (1) Synthesis of invasion literature within passenger-driver-backseat driver frameworks; (2) hydrochory investigation of water as a dispersal mechanism for invasion; (3) ecohydrology investigation of inundation effects on seed viability of L. -

Cash Thesis Final.Pdf (4.432Mb)
Evaluating Remote Sensing and Prescribed Fire Methods to Aid in the Restoration of Bottomland Hardwood Forests Invaded by Chinese Privet (Ligustrum sinense) by James Scollin Cash A thesis submitted to the Graduate Faculty of Auburn University in partial fulfillment of the requirements for the Degree of Master of Science Auburn, Alabama August 3, 2019 Approved by Christopher Anderson, Chair, Associate Professor of Wetland Ecology William Gulsby, Assistant Professor of Wildlife Management Luke Marzen, Professor of Geosciences John Kush, Research Fellow Abstract Chinese privet (Ligustrum sinense Lour.) is an invasive shrub with a wide distribution outside its native Southeast Asia range. It is particularly common and problematic in the southeastern U.S., where researchers have documented negative impacts to woody and herbaceous plant communities, which in turn likely affects wildlife habitat quality. This research project evaluated two management tools that could assist land managers in efforts to restore bottomland hardwood forests that have been invaded by L. sinense. Our first objective was to evaluate whether free multispectral satellite imagery and free simple-to-use software could be used to map L. sinense invasions and aid in planning and budgeting restoration projects. We found that the Semi-Automatic Classification Plugin within QGIS was effective at detecting L. sinense, particularly when using late dormant season Sentinel 2 imagery. Our second objective was to evaluate whether prescribed fire could reliably move through bottomland hardwood forests and reduce L. sinense slash following cutting operations to improve the efficiency of follow-up treatments on re-sprouts. We found that stand composition had the most important effect on fire spread (plots with greater proportions of tree species with flammable leaf traits tended to burn best), but that the fires were only successful in a limited number of cases. -

Obtuse-Leaved Privet, Border Privet Ligustrum Obtusifolium Siebold & Zucc
Obtuse-leaved privet, Border privet Ligustrum obtusifolium Siebold & Zucc. Olive Family (Oleaceae) DESCRIPTION Obtuse-leaved privet is a deciduous shrub that is multi- stemmed from the base, with spreading, twiggy, branches. It that has become extensively naturalized in disturbed woods and stream valleys. Several other species, Amur privet (L. amurense), California privet (L. ovalifolium), and common or European privet (L. vulgare) are also present as naturalized species in Pennsylvania; however, obtuse-leaved privet is the most common and invasive of these in our area. fruiting stem Height - This shrub grows 10–12 feet tall and 8–10 feet wide. Stem - Branches are slender, light gray, and exhibit an opposite arrangement along the stem. Young stems are hairy. Leaves - Leaves are opposite, firm, smooth, and 1–2 inches long, tapering slightly to a blunt tip and base. Summer leaf color is dark green, becoming purplish in the fall. Flowers - The small white flowers are in clusters at the ends of the branches; they are tubular with 4 spreading petal lobes, and about ⅓ inch long. Blooming occurs in June. Fruit and seed - The small (⅛ inch), black fruits are produced in clusters at the ends of branches; they mature in September and often persist on the plant all winter. Roots - Sprouting can occur from the roots when the tops are cut back severely. flowering stem DISTRIBUTION AND HABITAT Obtuse-leaved privet is native to Japan; it was introduced as a landscape plant in 1860. It has subsequently become naturalized from New Hampshire to North Carolina and west to Michigan, Indiana, and Tennessee. It is present throughout the southern half of Pennsylvania, and at scattered sites elsewhere in the state. -
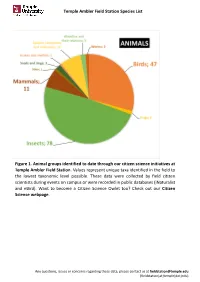
Temple Ambler Field Station Species List Figure 1. Animal Groups Identified to Date Through Our Citizen Science Initiatives at T
Temple Ambler Field Station Species List Figure 1. Animal groups identified to date through our citizen science initiatives at Temple Ambler Field Station. Values represent unique taxa identified in the field to the lowest taxonomic level possible. These data were collected by field citizen scientists during events on campus or were recorded in public databases (iNaturalist and eBird). Want to become a Citizen Science Owlet too? Check out our Citizen Science webpage. Any questions, issues or concerns regarding these data, please contact us at [email protected] (fieldstation[at}temple[dot]edu) Temple Ambler Field Station Species List Figure 2. Plant diversity identified to date in the natural environments and designed gardens of the Temple Ambler Field Station and Ambler Arboretum. These values represent unique taxa identified to the lowest taxonomic level possible. Highlighted are 14 of the 116 flowering plant families present that include 524 taxonomic groups. A full list can be found in our species database. Cultivated specimens in our Greenhouse were not included here. Any questions, issues or concerns regarding these data, please contact us at [email protected] (fieldstation[at}temple[dot]edu) Temple Ambler Field Station Species List database_title Temple Ambler Field Station Species List last_update 22October2020 description This database includes all species identified to their lowest taxonomic level possible in the natural environments and designed gardens on the Temple Ambler campus. These are occurrence records and each taxon is only entered once. This is an occurrence record, not an abundance record. IDs were performed by senior scientists and specialists, as well as citizen scientists visiting campus. -

A Jósika-Orgona (Syringa Josikaea Jacq
Doktori (Ph.D.) értekezés A JÓSIKA-ORGONA (SYRINGA JOSIKAEA JACQ. FIL. EX RCHB.) AKTUÁLIS ÉS TÖRTÉNETI BIOGEOGRÁFIÁJA Lendvay Bertalan Kertészettudományi Doktori Iskola Budapesti Corvinus Egyetem Genetika és Növénynemesítés Tanszék Budapest 2014 A doktori iskola megnevezése: Kertészettudományi Doktori Iskola tudományága: Növénytermesztési és kertészeti tudományok vezetője: Dr. Tóth Magdolna egyetemi tanár, DSc Budapesti Corvinus Egyetem, Kertészettudományi Kar, Gyümölcstermő Növények Tanszék Témavezetők: Dr. Höhn Mária egyetemi docens, CSc Budapesti Corvinus Egyetem, Kertészettudományi Kar, Növénytani Tanszék és Soroksári Botanikus Kert Dr. Pedryc Andrzej egyetemi tanár, DSc Budapesti Corvinus Egyetem, Kertészettudományi Kar, Genetika és Növénynemesítés Tanszék A jelölt a Budapesti Corvinus Egyetem Doktori Szabályzatában előírt valamennyi feltételnek eleget tett, az értekezés műhelyvitájában elhangzott észrevételeket és javaslatokat az értekezés átdolgozásakor figyelembe vette, azért az értekezés védési eljárásra bocsátható. ……………………………… ……………………………… Dr. Tóth Magdolna Dr. Höhn Mária ……………………………… Dr. Pedryc Andrzej A doktori iskola vezetőjének A témavezetők jóváhagyása jóváhagyása 2 A Budapesti Corvinus Egyetem Élettudományi Területi Doktori Tanácsának 2014.03.18-i határozatában a nyilvános vita lefolytatására az alábbi bíráló Bizottságot jelölte ki: BÍRÁLÓ BIZOTTSÁG: Elnöke Palkovics László, DSc Tagjai Bisztray György, PhD Deák Tamás, PhD Csiky János, PhD Pócs Tamás, MHAS Opponensek Halász Júlia, PhD Kovács J. Attila, CSc Titkár Deák -

Vascular Flora of the Yew Mountain Center, Pocahontas County, West Virginia
Proceedings of the West Virginia Academy of Science Original Research Paper Vascular Flora of the Yew Mountain Center, Pocahontas County, West Virginia Michael J. Baranski 1 1Department of Biology, Catawba College, Salisbury, NC, USA Article history Abstract: A baseline floristic study was conducted on the 500 acre Yew Received: 8 March 2021 Mountain Center (YMC) in Pocahontas County, West Virginia. The YMC Revised: 30 April 2021 is a non-profit educational center providing programs emphasizing Accepted: 3 May 2021 Appalachian ecology, culture, and arts, with one of its goals being the development of field research and study opportunities for outside *Corresponding Author: researchers and students. Field work conducted in 2019-2021 resulted in Michael J. Baranski, Catawba 383 recorded vascular plant taxa, with 25 new additions to the known College, Salisbury, NC, USA; Pocahontas County flora (possibly as many as 83 total new taxa have been Email: found by the author in the county). The majority of the new additions were [email protected] not unexpected. Two taxa (Juglans cinerea and Prunus alleghaniensis var. alleghaniensis) are significant occurrences tracked by the West Virginia Natural Heritage Program. Approximately 85% of the taxa are historical natives, as compared to 75% native flora for West Virginia as a whole. The primarily forested YMC harbors a diverse assemblage of Mixed Mesophytic, Mixed Hardwood and Appalachian Oak forest communities that is relatively free of exotic invasive and noxious weed infestations. The study demonstrates how a moderately-intensive examination of a small area can result in a significant increase in floristic knowledge of a region. Keywords: Yew Mountain Center, Pocahontas County, floristics.