Mitochondrial D-Loop Variation, Coat Colour and Sex Identification of Late
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Gravimetry for Geothermal Exploration
GRAVIMETRY FOR GEOTHERMAL EXPLORATION METHODOLOGY, COMPUTER PROGRAMS AND TWO CASE STUDIES IN THE SWISS MOLASSE BASIN Thesis defended on January 8, 2015 at the FACULTY OF SCIENCES INSTITUTE OF HYDROGEOLOGY AND GEOTHERMICS (CHYN) UNIVERSITY OF NEUCHÂTEL for the degree of DOCTOR OF NATURAL SCIENCES presented by PIERRICK ALTWEGG Accepted by the following Jury: PROF. PHILIPPE RENARD PROF. STEPHEN ANDREW MILLER PROF. GUILLAUME CAUMON PROF. JON MOSAR DR. EVA SCHILL DR. FRANÇOIS-DAVID VUATAZ DR. PIER-VITTORIO RADOGNA To my beloved wife and children, Alexandrine, Elanor and Aurèle. Acknowledgments A PhD thesis is something unique in the life of a scientist but is not a project that one can carry alone. In this respect, many individuals, institutions and societies contributed to it and I want to gratefully thank them. First, I owe much to Dr. François-David Vuataz who introduced me to the world of geothermal energy, saved me countless hours by being a living database and for her active participation in the project. I also would like to thank Prof. Eva Schill who made this project possible. A special thanks to Prof. Philippe Renard who agreed to supervise this work after three years and for his availability and his advice. I also want to thank Prof. Daniel Hunkeler and Prof. Pierre Perrochet for their support. My gratitude to all the members of the jury for their availability and their input after the critical review of this PhD thesis. I would like to especially thank Prof. Jon Mosar of the University of Fribourg for his time and input as well as for teaching me how to avoid formal errors in this work. -

Characterization of Bronze Casting in Insula 56, in Roman Aventicum
CHARACTERIZATION OF BRONZE CASTING IN INSULA 56, IN ROMAN AVENTICUM by JESSICA WHEAT COOK (Under the Direction of Ervan Garrison) ABSTRACT This study characterizes metallurgical wastes (slag) recovered from a bronze workshop during excavations in 1997-1998 at the Roman provincial city of Aventicum, located in western Switzerland, then Germania Superior. Previous research has identified the workshop as one devoted to the production of large leaded bronze statuary. The slag assemblage analyzed in this study is unusual due to the absence of ceramic crucible fragments normally associated with the production of copper alloys. Instead, this workshop is hypothesized to have used iron crucibles. Electron Microprobe Analysis (EMPA) and X-Ray Diffraction (XRD) were used to analyze the chemical composition of the slags in contrast to previous bulk analysis studies. Modern metal casting methods were observed for analogues. The data show that the slag reached temperatures between 1350°-1400° C and chemical components indicated the use of sand and lime fluxes. Evidence of high temperatures is proposed as one chemical fingerprint that can be used to re-assess slag for the presence of iron crucibles in other Roman bronze workshops. INDEX WORDS: Aventicum, Classical Bronzes, EMPA, XRD, Roman Metallurgy CHARACTERIZATION OF BRONZE CASTING IN INSULA 56, IN ROMAN AVENTICUM by JESSICA WHEAT COOK AB History, Anthropology, The University of Georgia, 2003 A Thesis Submitted to the Graduate Faculty of The University of Georgia in Partial Fulfillment of the Requirements for the Degree MASTER OF SCIENCE ATHENS, GEORGIA 2008 © 2008 Jessica Wheat Cook All Rights Reserved CHARACTERIZATION OF BRONZE CASTING IN INSULA 56, IN ROMAN AVENTICUM by JESSICA WHEAT COOK Major Professor: Ervan Garrison Committee: Samuel Swanson Douglas Crowe Electronic Version Approved: Maureen Grasso Dean of the Graduate School The University of Georgia May 2008 iv ACKNOWLEDGMENTS The author wishes to thank Drs. -
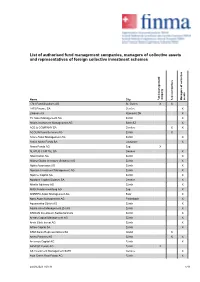
List of Authorised Fund Management Companies, Managers of Collective Assets and Representatives of Foreign Collective Investment Schemes
List of authorised fund management companies, managers of collective assets and representatives of foreign collective investment schemes Name City Fund management company Representatives Managers of collective assets 1741 Fund Solutions AG St. Gallen X X 1875 Finance SA Genève X 2Xideas AG Küsnacht ZH X 3V Asset Management AG Zürich X Abaris Investment Management AG Bäch SZ X ACE & COMPANY SA Genève X X ACOLIN Fund Services AG Zürich X Acoro Asset Management AG Zürich X Active Niche Funds SA Lausanne X Akara Funds AG Zug X ALATUS CAPITAL SA Genève X Albin Kistler AG Zürich X Allianz Global Investors (Schweiz) AG Zürich X Alpha Associates AG Zürich X Alpinum Investment Management AG Zürich X Alprime Capital AG Zürich X Alpstone Capital (Suisse) SA Genève X Altrafin Advisory AG Zürich X AMG Fondsverwaltung AG Zug X ANREPA Asset Management AG Baar X Apex Asset Management AG Freienbach X Aquamarine Zürich AG Zürich X Aquila Asset Management ZH AG Zürich X ARDIAN Investment Switzerland AG Zürich X Aretas Capital Management AG Zürich X Arete Ethik Invest AG Zürich X Arfina Capital SA Zürich X ARM Swiss Representatives SA Gland X Artico Partners AG Zürich X X Arvernus Capital AG Zürich X ARVEST Funds AG Zürich X AS Investment Management SARL Genève X Asia Green Real Estate AG Zürich X 24.09.2021 / 03:31 1/11 List of authorised fund management companies, managers of collective assets and representatives of foreign collective investment schemes Name City Fund management company Representatives Managers of collective assets Asteria Investment Managers SA Genève X Atlanticomnium SA Genève X AtonRâ Partners SA Genève X AUTONOMY CAPITAL RESEARCH TWO LIMITED, (St. -

The Roman Antiquities of Switzerland
Archaeological Journal ISSN: 0066-5983 (Print) 2373-2288 (Online) Journal homepage: http://www.tandfonline.com/loi/raij20 The Roman Antiquities of Switzerland By Bunnell Lewis M.A., F.S.A. To cite this article: By Bunnell Lewis M.A., F.S.A. (1885) The Roman Antiquities of Switzerland, Archaeological Journal, 42:1, 171-214, DOI: 10.1080/00665983.1885.10852174 To link to this article: http://dx.doi.org/10.1080/00665983.1885.10852174 Published online: 15 Jul 2014. Submit your article to this journal View related articles Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=raij20 Download by: [University of California Santa Barbara] Date: 18 June 2016, At: 04:49 THE ROMAN ANTIQUITIES OF SWITZERLAND. By BUNNELL LEWIS, M.A., F.S.A. Many persons, well-informed in other respects, think that there are no Boman antiquities in Switzerland. This mistake results from various causes. Most people travel there to enjoy the scenery, and recruit their health. The Bomans have not left behind them in that country vast monuments of their power, like the temples, theatres and aqueducts, which in regions farther south are still to be seen ; but, speaking generally, we must be content with smaller objects stored in museums, sometimes unprovided with catalogues.1 Moreover, no English writer, as far as I know, has discussed this subject at any length; attention has been directed almost exclusively to pie-historic remains made known by Dr. Keller's book on Bfahlbauten (lake- dwellings), of which an excellent translation has been published.2 However, I hope to show that the classical antiquities of Switzerland, though inferior to those of some other countries, ought not to be passed over with contemptuous neglect, and that they deserve study quite as much as similar relics of the olden time in Britain, 1 A very good account of the Collections 2 Dr. -

List of Swiss Cultural Goods: Archaeological Objects
List of Swiss cultural goods: archaeological objects A selection compiled by the Conference of Swiss Canton Archaeologists by order of the Federal Office of Culture FOC Konferenz schweizerischer Kantonsarchäologinnen und Kantonsarchäologen KSKA Conférence Suisse des Archéologues Cantonaux CSAC Conferenza Svizzera degli Archeologi Cantonali CSAC Categories of Swiss cultural goods I. Stone A. Architectural elements: made of granite, sandstone, marble and other types of stone. Capitals, window embrasures, mosaics etc. Approximate date: 50 BC – AD 1500. B. Inscriptions: on various types of stone. Altars, tombstones, honorary inscriptions etc. Approximate date: 50 BC – AD 800. C. Reliefs: on limestone and other types of stone. Stone reliefs, tomb- stone reliefs, decorative elements etc. Approximate date: mainly 50 BC – AD 800. D. Sculptures/statues: made of limestone, marble and other types of stone. Busts, statuettes, burial ornaments etc. Approximate date: mainly 50 BC – AD 800. E. Tools/implements: made of flint and other types of stone. Various tools such as knife and dagger blades, axes and implements for crafts etc. Approximate date: 130 000 BC – AD 800. F. Weapons: made of shale, flint, limestone, sandstone and other types of stone. Arrowheads, armguards, cannonballs etc. Approxi- mate date: 10 000 BC – AD 1500. G. Jewellery/accessories: made of various types of stone. Pendants, beads, finger ring inlays etc. Approximate date: mainly 2800 BC – AD 800. II. Metal A. Statues /statuettes / made of non-ferrous metal, rarely from precious metal. busts: Depictions of animals, humans and deities, portrait busts etc. Approximate date: 50 BC – AD 800. B. Vessels: made of non-ferrous metal, rarely from precious metal and iron. -

Calcareous Nannofossils, Ammonites
De Kaenel et al. Swiss J Palaeontol (2020) 139:6 https://doi.org/10.1186/s13358-020-00209-5 Swiss Journal of Palaeontology RESEARCH ARTICLE Open Access New biostratigraphical data (calcareous nannofossils, ammonites) and Early to Late Barremian transition in the Urgonien Jaune facies and Marnes de la Russille complex of the Swiss Jura Mountains Eric De Kaenel1*, Pierre‑Olivier Mojon2 and Antoine Pictet3 Abstract In the central Jura Mountains (Western Switzerland), the Urgonien Jaune (UJ) facies with the Marnes de la Russille beds (MRu) have provided very rich nannoforas associated with very rare Tethyan ammonites. A late Early Barremian nannofora of the Mid‑Barremian Event (MBE, following a regional tectonic event of an earliest Barremian synsedimentary tectonic crisis) was found in MRu of the lower UJ and includes 42 genera with 90 species. Among them, Biscutum jurensis De Kaenel, n. sp., Flabellites eclepensensis De Kaenel, n. sp., Palaeopontosphaera giraudii De Kaenel, n. sp., Rhagodiscus buisensis De Kaenel, n. sp., and Vagalapilla rutledgei De Kaenel, n. sp., are recognized as fve new species. This nannofora is a mixture of Boreal and Tethyan taxa with 20 nannofossil markers (Assipetra terebrodentarius, Broinsonia galloisii, Calcicalathina oblongata, Cyclagelosphaera rotaclypeata, Diloma placinum, Ethmorhabdus hauterivianus, Flabellites eclepensensis, Gorkaea pseudoanthophorus, Nannoconus abundans, Nannoconus pseudoseptentrionalis, Palaeopontosphaera giraudii, Palaeopontosphaera pinnata, Placozygus howei, Placozygus reticulatus, Reinhardtites scutula, Rhagodiscus buisensis, Rhagodiscus eboracensis, Tegulalithus septentrionalis, Tubodiscus jurapelagicus, Zeugrhabdotus moulladei) indicating very precisely the nannofossil Zones LK19 (Boreal)–NC5D (Tethyan) as well as the Elegans (Boreal) and Moutonianum (Tethyan) ammonite Zones of the latest Early Barremian. The ammonites in the basal UJ facies of Early Barremian age are reworked Lyticoceras claveli (Nodosoplicatum Zone, Early Hauterivian) and reworked Cruasiceras cf. -

Découverte Celtique Exceptionnelle En 2006 Le «Sanctuaire»
Découverte celtique exceptionnelle en 2006 Le «sanctuaire» Le site au sommet du Mormont, en cours de fouille. helvète du Mormont par Gilbert Kaenel et Alors que la communauté scientifique s’apprêtait à commémo- Denis Weidmann rer le cent cinquantième anniversaire de la découverte de La Tène en novembre 1857, le canton de Vaud mettait au jour en 2006, au sommet du Mormont, de manière inattendue, un site appelé à devenir une référence, au même titre que le site épo- nyme du Second âge du fer européen; référence non seule- ment pour l’archéologie des Helvètes du Plateau suisse, entre la fin du IIe et le début du Ier siècle av. J.-C., mais pour l’étu- de des pratiques rituelles et cultuelles des Celtes en général. 16 NIKE-Bulletin 4 /2007 NIKE-Bulletin 4 /2007 17 La Section de l’archéologie cantonale Venoge, au sud: le Mormont sépare ainsi les vaudoise a dû gérer une nouvelle interven- bassins versants du Rhin et du Rhône. Fosses et puits à offrandes tion nécessitant la mise en œuvre de moyens L’occupation de la fin de l’âge du Fer au sein d’un vaste ensemble à importants, affectés à l’organisation d’une s’est développée dans le fond d'un secteur vocation cultuelle fouille préventive de grande ampleur. Le légèrement encaissé, encadré par des fail- Quelque 300 structures (fosses, trous de po- Musée cantonal d’archéologie et d’histoire les, et proche du sommet (604 m). La sur- teau, foyers, blocs de signalisation, petits de Lausanne de son côté, responsable de la face rocheuse, retravaillée par l’érosion et fossés) ont été mises au jour; parmi ces der- conservation des vestiges exhumés, a dû en particulier par l’activité glaciaire qua- nières, plus de 260 fosses sont interprétées adapter l’activité de son laboratoire à l’arri- ternaire, présente un aspect moutonné avec comme de véritables puits à offrandes. -

Switzerland Buried by Glaciers During the Ice Age, Switzerland Was Colonized Around 15,000 Years Ago
TOP HISTORY DESTINATIONS: Switzerland Buried by glaciers during the Ice Age, Switzerland was colonized around 15,000 years ago. Discover the pasts and struggles with the Celtic Tribes, Helvetians, Greeks, and Romans. Explore the Middle Ages origins of the split between German and French languages with the Burgundians and Alamannen. Don’t miss this scenic land whose medieval roots, chocolate, cheese, banking system, watches, and alpine mountain areas make it one of the happiest places on earth and makes it a wonderful destination for student history trips. EDUCATIONAL HISTORY OPPORTUNITIES: • Experience the Life of the Gladiators • Visit a Gorgeous Traditional Resort Town • See the Swiss Grand Canyon, a Deep with Interactive Combat Training and in the Mountainous Bernese Gorge Created Over 10,000 Years by the Gladiator School • Explore the Largest Town and capital of Ice Age Rhine Glacier • Learn How the Swiss Confederation was Roman Switzerland • See How the Inhabitants of Zermatt Able to Defeat the Army of Knights • Visit a Well-Preserved Former Lived in the 19th Century • Go Back in Time in the Halls and Rooms Benedictine Monastery • Visit a Geneva Cathedral Where John of the Castle of Chillon • Explore the Best-Preserved Fossils that Calvin Choose to Preach • Tour the Headquarters of the League of Date Back 230 Million Years to the • Explore the Oldest Wooden Bridge in Nations Triassic Period Switzerland Dating Back to the Middle • See the Saddest Stone in the World • Learn How people lived way back during of the 14th Century Commemorating -

De L'âge Du Fer À L'usage Du Verre
Cahiers d'archéologie romande 151 De l'âge du Fer à l'usage du verre Mélanges offerts à Gilbert Kaenel, dit «Auguste», à l'occasion de son 65e anniversaire Textes réunis par Jérôme Bullinger, Pierre Crotti et Claire Huguenin Lausanne 2014 Sommaire Avant-propos Jérôme BULLINGER, Pierre CROTTI et Claire HUGUENIN 9 Préface d'un coéditeur Denis WEIDMANN 11 «Rendez-vous à la Couronne !» Suzanne et Daniel FIBBI-AEPPLI 13 A propos d'une formule orale usuelle en contexte vaudois : approche analytique, interprétative et tout le commerce Laurent FLUTSCH 15 Clin d'oeil sur la portée du temps... Daniel PAUNIER 21 Frontière... quelle frontière ? Gilbert Kaenel et la valorisation hors de Suisse de la recherche sur l'âge du Fer chez les Helvètes : l'archéologie à saute-frontière Marie-Jeanne ROULIÈRE-LAMBERT, Philippe BARRAL, Philippe CURDY, Cynthia DUNNING-THIERSTEIN, 25 Olivier BUCHSENSCHUTZ, Jean-Loup FLOUEST et François SCHIFFERDECKER De Naef à Auguste : La Tène et les Vaudois Marc-Antoine KAESER et Gianna REGINELLI SERVAIS 37 Les archives en musées et en bibliothèques Gilbert COUTAZ 43 De Lausanne à Lattes : réflexions sur les expositions d'archéologie Lionel PERNET 49 L'église-musée. Réflexions sur l'exposition des vestiges archéologiques dans les églises de Suisse romande au 20e siècle Dave LUTH! en collaboration avec Claire HUGUENIN 59 De la fouille aux restitutions en Protohistoire : l'exemple du sanctuaire de Gournay-sur-Aronde Stephan FICHTL 69 Bien culturel et valeur économique : un conflit devenu omniprésent Marc-André HALDIMANN 77 Processus -

Archaeological Journal Notice of a Die for Striking
This article was downloaded by: [Northwestern University] On: 24 January 2015, At: 14:59 Publisher: Routledge Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Archaeological Journal Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/raij20 Notice of a Die for Striking Helvetian or Gaulish Gold Coins; Found at Avenches in Switzerland Dr. Ferdinand Keller President of the Society of Antiquaries of Zürich. Hon. Corr. Member of the Archaeological Institute Published online: 10 Jul 2014. To cite this article: Dr. Ferdinand Keller President of the Society of Antiquaries of Zürich. Hon. Corr. Member of the Archaeological Institute (1862) Notice of a Die for Striking Helvetian or Gaulish Gold Coins; Found at Avenches in Switzerland, Archaeological Journal, 19:1, 253-258, DOI: 10.1080/00665983.1862.10851220 To link to this article: http://dx.doi.org/10.1080/00665983.1862.10851220 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. -

Arch, Die Römerstrasse
Erziehungsdirektion des Kantons Bern Direction de l’instruction publique du canton de Berne Amt für Kultur | Office de la culture Archäologischer Dienst des Kantons Bern Service archéologique du canton de Berne Postfach 5233, 3001 Bern Telefon 031 633 98 22 5 1 2 3 4 [email protected] www.be.ch/archaeologie Die römische Strasse besteht aus: 1 einem ursprünglich 6 m breiten, im Querschnitt linsen förmigen, gebänderten Kieskörper, der aus mehreren, immer wieder erneuerten Fahrbahnen zusammengesetzt ist. ARCH 2 Für die erste Fahrbahn wurde der Untergrund mit einer Stein setzung verfestigt. Die Römerstrasse 3 Auf den einzelnen Fahrbahnen zeichnen sich die Karrengeleise ab. Meist lassen sich vier Hauptgeleise ausmachen. Sie gehören zu zwei 1,4 m breiten Wagenspuren, deren innere Geleise sich überkreuzen. 4 Bergseits begleitet ein aus Kieseln gebauter Geh- oder Viehweg (iter oder actus) die Fahrbahn. Nützliche Hinweise: Ein Ausschnitt der Römerstrasse ist präpariert und in einem Schauraum beim Römer Café, Römerstrasse West 24 in Arch zu 5 Zwei Spitzgräben begleiten die Fahrbahn beidseitig in etwa 6 m Ent- besichtigen. fernung. Der talseitige Graben liegt ausserhalb des Schauraums. Gestaltung: Archäologischer Dienst des Kantons Bern, René Bacher (Text). Literatur: René Bacher und K. Ramseyer, Arch und Büren a. A. 1991. Zur Römerstrasse zwischen Petinesca und Salodurum. Archäologie im Kanton Bern 3B. Bern 1994, 375–391. Titelbild: Schauraum neben dem Römer Café. � Blick auf die freipräparier- ten Fahrbahnen im Schau- Bildnachweis: Archäologischer Dienst des Kantons Bern. raum. © 2014 ADB / René Bacher (Text), Badri Redha (Fotos), Max Stöckli (Grafik). Archäologischer Dienst des Kantons Bern 8/2014 Service archéologique du canton de Berne Das Strassennetz bildet das Rückgrat des römischen Reiches. -

Her Fabulous Story!
AVENTICUM HER FABULOUS STORY! 3rd floor Copyright © Site et Musée romains d’Avenches October 2019 Translation Natasha Hathaway www.archeo-access.ch Scenography www.wapico.ch Multimedia www.pointprod.ch Aventicum: a people, a territory The story of Aventicum begins a long time Amongst the inhabitants were members before the Roman conquest. In the middle of the imperial family, such as the father of the 2nd century BCE, it was already of Vespasian and his son Titus, prominent a major Gallic urban centre. The local families, lower class citizens, doctors, aristocracy minted coins and traded with traders, craftsmen, free men, freedmen Mediterranean regions. and slaves. In 15 BCE, the territory of the Helvetii, Economic and political troubles marked from Lake Geneva to Lake Constance, went the end of the 3rd century and large areas under Roman control. Avenches became the of the city were abandoned. However, capital of the Helvetii and the town quickly the city became and remained an episcopal expanded. During the 2nd century CE, there see until the end of the 6th century CE. It were approximately 20,000 people living in survived only as a small-sized settlement in Avenches, most of them indigenous. the following centuries. In the 13th century, the “new town” was built on the hilltop. 4 5 Gallic origins Until recently, it was assumed that the city of Aventicum had been created ex nihilo soon after 15 BCE, when the Helvetii territory was integrated into the Roman Empire. Over the past few years, discoveries of Celtic remains west and south of the hill of Avenches have brought to light that a large urban centre already existed in the second half of the 2nd century BCE.