Reproductive Behavior, Development and Eye Regression in the Cave
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Preserving the Tree of Life of the Fish Family Cyprinidae in Africa in the Face of the Ongoing Extinction Crisis
Genome Preserving the tree of life of the fish family Cyprinidae in Africa in the face of the ongoing extinction crisis Journal: Genome Manuscript ID gen-2018-0023.R3 Manuscript Type: Article Date Submitted by the 02-Mar-2019 Author: Complete List of Authors: Adeoba, Mariam; University of Johannesburg Tesfamichael, Solomon; University of Johannesburg Yessoufou, Kowiyou; University of Johannesburg Conservation, African freshwater Ecosystems, IUCN Red List, EDGE, DNA Keyword: Draft barcoding Is the invited manuscript for consideration in a Special 7th International Barcode of Life Issue? : https://mc06.manuscriptcentral.com/genome-pubs Page 1 of 42 Genome Preserving the tree of life of the fish family Cyprinidae in Africa in the face of the ongoing extinction crisis Mariam Salami1, Solomon Tesfamichael2, Kowiyou Yessoufou2 1Department of Zoology, University of Johannesburg, Kingsway Campus, PO Box 524, Auckland Park 2006, South Africa 2Department of Geography, Environmental Management and Energy studies, University of Johannesburg, Kingsway Campus, PO Box 524, Auckland Park 2006, South Africa *Corresponding author: Kowiyou Yessoufou [email protected] Draft 1 https://mc06.manuscriptcentral.com/genome-pubs Genome Page 2 of 42 Abstract Our understanding of how the phylogenetic tree of fishes might be affected by the ongoing extinction risk is poor. This is due to the unavailability of comprehensive DNA data, especially for many African lineages. In addition, the ongoing taxonomic confusion within some lineages, e.g. Cyprinidae, makes it difficult to contribute to the debate on how the fish tree of life might be shaped by extinction. Here, we combine COI sequences and taxonomic information to assemble a fully sampled phylogeny of the African Cyprinidae and investigate whether we might lose more phylogenetic diversity (PD) than expected if currently-threatened species go extinct. -

Checklist of Fish and Invertebrates Listed in the CITES Appendices
JOINTS NATURE \=^ CONSERVATION COMMITTEE Checklist of fish and mvertebrates Usted in the CITES appendices JNCC REPORT (SSN0963-«OStl JOINT NATURE CONSERVATION COMMITTEE Report distribution Report Number: No. 238 Contract Number/JNCC project number: F7 1-12-332 Date received: 9 June 1995 Report tide: Checklist of fish and invertebrates listed in the CITES appendices Contract tide: Revised Checklists of CITES species database Contractor: World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge, CB3 ODL Comments: A further fish and invertebrate edition in the Checklist series begun by NCC in 1979, revised and brought up to date with current CITES listings Restrictions: Distribution: JNCC report collection 2 copies Nature Conservancy Council for England, HQ, Library 1 copy Scottish Natural Heritage, HQ, Library 1 copy Countryside Council for Wales, HQ, Library 1 copy A T Smail, Copyright Libraries Agent, 100 Euston Road, London, NWl 2HQ 5 copies British Library, Legal Deposit Office, Boston Spa, Wetherby, West Yorkshire, LS23 7BQ 1 copy Chadwick-Healey Ltd, Cambridge Place, Cambridge, CB2 INR 1 copy BIOSIS UK, Garforth House, 54 Michlegate, York, YOl ILF 1 copy CITES Management and Scientific Authorities of EC Member States total 30 copies CITES Authorities, UK Dependencies total 13 copies CITES Secretariat 5 copies CITES Animals Committee chairman 1 copy European Commission DG Xl/D/2 1 copy World Conservation Monitoring Centre 20 copies TRAFFIC International 5 copies Animal Quarantine Station, Heathrow 1 copy Department of the Environment (GWD) 5 copies Foreign & Commonwealth Office (ESED) 1 copy HM Customs & Excise 3 copies M Bradley Taylor (ACPO) 1 copy ^\(\\ Joint Nature Conservation Committee Report No. -

A Guide to the Parasites of African Freshwater Fishes
A Guide to the Parasites of African Freshwater Fishes Edited by T. Scholz, M.P.M. Vanhove, N. Smit, Z. Jayasundera & M. Gelnar Volume 18 (2018) Chapter 2.1. FISH DIVERSITY AND ECOLOGY Martin REICHARD Diversity of fshes in Africa Fishes are the most taxonomically diverse group of vertebrates and Africa shares a large portion of this diversity. This is due to its rich geological history – being a part of Gondwana, it shares taxa with the Neotropical region, whereas recent close geographical affnity to Eurasia permitted faunal exchange with European and Asian taxa. At the same time, relative isolation and the complex climatic and geological history of Africa enabled major diversifcation within the continent. The taxonomic diversity of African freshwater fshes is associated with functional and ecological diversity. While freshwater habitats form a tiny fraction of the total surface of aquatic habitats compared with the marine environment, most teleost fsh diversity occurs in fresh waters. There are over 3,200 freshwater fsh species in Africa and it is likely several hundreds of species remain undescribed (Snoeks et al. 2011). This high diversity and endemism is likely mirrored in diversity and endemism of their parasites. African fsh diversity includes an ancient group of air-breathing lungfshes (Protopterus spp.). Other taxa are capable of breathing air and tolerate poor water quality, including several clariid catfshes (e.g., Clarias spp.; Fig. 2.1.1D) and anabantids (Ctenopoma spp.). Africa is also home to several bichir species (Polypterus spp.; Fig. 2.1.1A), an ancient fsh group endemic to Africa, and bonytongue Heterotis niloticus (Cuvier, 1829) (Osteoglossidae), a basal actinopterygian fsh. -

Volume 2. Animals
AC20 Doc. 8.5 Annex (English only/Seulement en anglais/Únicamente en inglés) REVIEW OF SIGNIFICANT TRADE ANALYSIS OF TRADE TRENDS WITH NOTES ON THE CONSERVATION STATUS OF SELECTED SPECIES Volume 2. Animals Prepared for the CITES Animals Committee, CITES Secretariat by the United Nations Environment Programme World Conservation Monitoring Centre JANUARY 2004 AC20 Doc. 8.5 – p. 3 Prepared and produced by: UNEP World Conservation Monitoring Centre, Cambridge, UK UNEP WORLD CONSERVATION MONITORING CENTRE (UNEP-WCMC) www.unep-wcmc.org The UNEP World Conservation Monitoring Centre is the biodiversity assessment and policy implementation arm of the United Nations Environment Programme, the world’s foremost intergovernmental environmental organisation. UNEP-WCMC aims to help decision-makers recognise the value of biodiversity to people everywhere, and to apply this knowledge to all that they do. The Centre’s challenge is to transform complex data into policy-relevant information, to build tools and systems for analysis and integration, and to support the needs of nations and the international community as they engage in joint programmes of action. UNEP-WCMC provides objective, scientifically rigorous products and services that include ecosystem assessments, support for implementation of environmental agreements, regional and global biodiversity information, research on threats and impacts, and development of future scenarios for the living world. Prepared for: The CITES Secretariat, Geneva A contribution to UNEP - The United Nations Environment Programme Printed by: UNEP World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge CB3 0DL, UK © Copyright: UNEP World Conservation Monitoring Centre/CITES Secretariat The contents of this report do not necessarily reflect the views or policies of UNEP or contributory organisations. -

Etude Comparã©E Du Comportement
Int. J. Speleol. 4 (1972), pp. 139-169. Etude comparee du comportement alimentaire de deux poissons cavernicoles (Anoptichthys jordani Hubbs et Innes et Caecobarbus geertsi Blgr.l par Georges THINES et Nicole WISSOCQ* INTRODUCTION Dans un travail preliminaire pam dans ce meme Journal, Thines, Soffie et Vandenbussche (1966) avaient procede a l'analyse du comportement alimentaire du poisson cavernicole Anoptichthys Gen. et des hybrides F 1 obtenus par croisement Astyanax x Anoptichthys, la premiere espece etant la forme ancestrale epigee de la seconde. Les resultats de ces recherches avaient mis en evidence les traits essentiels de la reaction de la forme cavernicole Anoptichthys a la stimulation chimique. Ces donnees avaient egalement permis de caracteriser la stereotypie d'exploration de ce poisson consecutive a la stimulation chimique initiale. Les auteurs precites avaient etabli que l'exploration alimentaire s'effectuait sous la forme d'une plongee vers Ie fond suivie d'une recherche active localisee autour du point d'impact de la nourriture sur Ie substrat. L'efficacite de saisie, comparee a celIe de la forme ancestrale Astyanax, est tres deficitaire. De plus, la plongee vers Ie fond peut etre declenchee par des stimuli perturbants de nature mecanique. L'ensemble de ces observations avait ete soumis a la discussion et avait amene a conclure que Ie comportement alimentaire, outre les deficits evidents qu'il accuse sur Ie plan de l'efficacite spatio-temporelle de saisie, intervenait a titre substitutif dans des situations declenchant des comportements phobiques typiques chez la forme epigee Astyanax. POSITION DU PROBLEME Si I'on se refere a la classification de Vandel (1964), l'Anoptichthys est un cavernicole recent, tandis que Ie Caecobarbus est un cavernicole ancien. -
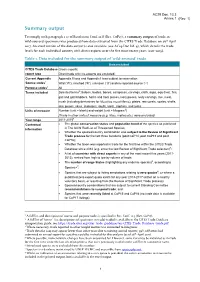
Summary Output
AC29 Doc. 13.3 Annex 1 Summary output To comply with paragraph 1 a) of Resolution Conf. 12.8 (Rev. CoP17), a summary output of trade in wild-sourced specimens was produced from data extracted from the CITES Trade Database on 26th April 2017. An excel version of the data output is also available (see AC29 Doc Inf. 4), which details the trade levels for each individual country with direct exports over the five most recent years (2011-2015). Table 1. Data included for the summary output of ‘wild-sourced’ trade Data included CITES Trade Database Gross exports; report type Direct trade only (re-exports are excluded) Current Appendix Appendix II taxa and Appendix I taxa subject to reservation Source codes1 Wild (‘W’), ranched (‘R’), unknown (‘U’) and no reported source (‘-’) Purpose codes1 All Terms included Selected terms2: baleen, bodies, bones, carapaces, carvings, cloth, eggs, egg (live), fins, gall and gall bladders, horns and horn pieces, ivory pieces, ivory carvings, live, meat, musk (including derivatives for Moschus moschiferus), plates, raw corals, scales, shells, skin pieces, skins, skeletons, skulls, teeth, trophies, and tusks. Units of measure Number (unit = blank) and weight (unit = kilogram3) [Trade in other units of measure (e.g. litres, metres etc.) were excluded] Year range 2011-20154 Contextual The global conservation status and population trend of the species as published information in The IUCN Red List of Threatened Species; Whether the species/country combination was subject to the Review of Significant Trade process for the last three iterations (post CoP14, post CoP15 and post CoP16); Whether the taxon was reported in trade for the first time within the CITES Trade Database since 2012 (e.g. -

Inventaire Rapide Des Zones Humides Représentatives En République Démocratique Du Congo
Inventaire Rapide des Zones Humides Représentatives en République Démocratique du Congo Editors and Major Contributors : Michele Thieme, Aurelie Shapiro, Alejandra Colom, Uli Schliewen, Nikolai Sindorf, Andre Kamdem Toham Supported by the Swiss Grant for Africa, Ramsar Convention Secretariat Table of Contents Executive Summary I. Introduction II. Approach and Methodology for Rapid Inventory of Wetland Priority Areas A. Aquatic Ecosystem Classification and Gap Analysis B. Human Use / Suitability Analysis C. Marxan Model D. Delineation and Prioritization of Wetland Areas E. Threats assessment, socio economic importance and key stakeholders Appendix I. Contributors Appendix II. List of Protected Areas used in the Gap Analysis Appendix III. Photos of Wetland Areas Appendix IV. List of Fish Species by Wetland Priority Area Executive Summary In 2002, the government of the Democratic Republic of Congo committed to increase its protected area coverage to 15% from just over 8%. WWF, RAMSAR, OSFAC and other partners supported the effort by convening an expert workshop in Kinshasa, DRC in November 2007, to undertake a country wide biodiversity assessment, including a rapid inventory and prioritization of the biodiversity value of the wetlands of the Democratic Republic of Congo. Several products were developed prior to the workshop to facilitate the work of the experts. These were: (1) a map classifying small sub‐basins in DRC by the predominant aquatic habitat types and (2) a map of human use/suitability. The aquatic habitat or ecosystem map was used as a basis for ensuring representation of all habitat types across the country and the suitability map provided a measure of the level of human impact. -

(Siluriformes, Loricariidae) of the Paraguay River Basin
COMPARATIVE A peer-reviewed open-access journal CompCytogenCytogenetic 10(4): 625–636data on (2016)Ancistrus sp. (Siluriformes, Loricariidae) of the Paraguay River basin... 625 doi: 10.3897/CompCytogen.v10i4.8532 RESEARCH ARTICLE Cytogenetics http://compcytogen.pensoft.net International Journal of Plant & Animal Cytogenetics, Karyosystematics, and Molecular Systematics Cytogenetic data on Ancistrus sp. (Siluriformes, Loricariidae) of the Paraguay River basin (MS) sheds light on intrageneric karyotype diversification Ana Camila Prizon1, Luciana Andreia Borin-Carvalho1, Daniel Pacheco Bruschi3, Marcos Otávio Ribeiro1, Ligia Magrinelli Barbosa1, Greicy Ellen de Brito Ferreira1, Andréa Cius1, Claudio Henrique Zawadzki2, Ana Luiza de Brito Portela-Castro1 1 Departamento de Biotecnologia, Genética e Biologia Celular, Universidade Estadual de Maringá, UEM, Avenida Colombo, 5790, 87020-900, Maringá, Paraná, Brazil 2 Departamento de Biologia/ Nupélia, Universidade Estadual de Maringá, UEM, Avenida Colombo, 5790, 87020-900, Maringá, Paraná, Brazil 3 Departamento de Genética, Universidade Federal do Paraná, UFPR, Centro Politécnico, Jardim Botânico, 81531-980, Curitiba, Paraná, Brazil Corresponding author: Ana Camila Prizon ([email protected]) Academic editor: A. Boron | Received 17 March 2016 | Accepted 17 October 2016 | Published 22 November 2016 http://zoobank.org/82711DD1-E2E4-41A0-8391-FA6CDD84FE24 Citation: Prizon AC, Borin-Carvalho LA, Bruschi DP, Ribeiro MO, Barbosa LM, Ferreira GEB, Cius A, Zawadzki CH, Portela-Castro ALB (2016) Cytogenetic data on Ancistrus sp. (Siluriformes, Loricariidae) of the Paraguay River basin (MS) sheds light on intrageneric karyotype diversification. Comparative Cytogenetics 10(4): 625–636. doi: 10.3897/CompCytogen.v10i4.8532 Abstract Ancistrus Kner, 1854 is a diverse catfish genus, currently comprising 66 valid species, but karyotype data were recorded for 33 species, although only ten have their taxonomic status defined. -

AC31 Doc. 13.4 A2
AC31 Doc.13.4 Annex 2 AC31 Doc. xx Annex X UNEP-WCMC Technical Report SELECTION OF SPECIES FOR INCLUSION IN THE REVIEW OF SIGNIFICANT TRADE FOLLOWING COP18: Extended Analysis AC31 Doc. 13.4 Annex 2 Selection of species for inclusion in the Review of Significant Trade following CoP18: Extended analysis Prepared for CITES Secretariat Published May 2020 Citation UNEP-WCMC. 2020. Selection of species for inclusion in the Review of Significant Trade following CoP18: Extended analysis. UNEP-WCMC, Cambridge. Copyright CITES Secretariat, 2020 The UN Environment Programme World Conservation Monitoring Centre (UNEP-WCMC) is a global Centre of excellence on biodiversity. The Centre operates as a collaboration between the UN Environment Programme and the UK-registered charity WCMC. Together we are confronting the global crisis facing nature. This publication may be reproduced for educational or non-profit purposes without special permission, provided acknowledgement to the source is made. Reuse of any figures is subject to permission from the original rights holders. No use of this publication may be made for resale or any other commercial purpose without permission in writing from the UN Environment Programme. Applications for permission, with a statement of purpose and extent of reproduction, should be sent to the Director, UNEP-WCMC, 219 Huntingdon Road, Cambridge, CB3 0DL, UK. The contents of this report do not necessarily reflect the views or policies of the UN Environment Programme, contributory organisations or editors. The designations employed and the presentations of material in this report do not imply the expression of any opinion whatsoever on the part of the UN Environment Programme or contributory organisations, editors or publishers concerning the legal status of any country, territory, city area or its authorities, or concerning the delimitation of its frontiers or boundaries or the designation of its name, frontiers or boundaries. -

Loricariidae)
UNIVERSIDADE FEDERAL DO PARÁ INSTITUTO DE CIÊNCIAS BIOLÓGICAS PÓS-GRADUAÇÃO EM ECOLOGIA AQUÁTICA E PESCA THAYANA AYRES ALVES MAPEAMENTO FÍSICO DE DNAs REPETITIVOS EM ESPÉCIES DA SUBFAMÍLIA HYPOSTOMINAE (LORICARIIDAE) Belém – PA 2016 THAYANA AYRES ALVES MAPEAMENTO FÍSICO DE DNAs REPETITIVOS EM ESPÉCIES DA SUBFAMÍLIA HYPOSTOMINAE (LORICARIIDAE) Dissertação apresentado ao Colegiado do Curso de Pós Graduação em Ecologia Aquática e Pesca, da Universidade Federal do Pará, como requisito para do título de mestre em Ecologia Aquática e Pesca. Orientadora: Profª. Drª. Renata Coelho Rodrigues Noronha. Belém – PA 2016 THAYANA AYRES ALVES MAPEAMENTO FÍSICO DE DNAs REPETITIVOS EM ESPÉCIES DA SUBFAMÍLIA HYPOSTOMINAE (LORICARIIDAE) Dissertação apresentado ao Colegiado do Curso de Pós Graduação em Ecologia Aquática e Pesca, da Universidade Federal do Pará, como requisito para do título de mestre em Ecologia Aquática e Pesca. Banca Examinadora: Profª. Drª. Renata Coelho Rodrigues Noronha (Presidente) Profº. Dr. Julio Cesar Pieczarka (membro) Profº. Dr. Pablo Suárez (membro) Profº. Dr. Leonardo dos Santos Sena (membro) Profº. Dr. Carlos Eduardo Matos Carvalho Bastos (suplente) Belém – PA 2016 THAYANA AYRES ALVES MAPEAMENTO FÍSICO DE DNAs REPETITIVOS EM ESPÉCIES DA SUBFAMÍLIA HYPOSTOMINAE (LORICARIIDAE) Dissertação apresentado ao Colegiado do Curso de Pós Graduação em Ecologia Aquática e Pesca, da Universidade Federal do Pará, Instituto de Ciências Biológicas. Banca Examinadora: _________________________________________ Profª. Drª. Renata Coelho Rodrigues Noronha (Presidente) Laboratório de Citogenética Animal da UFPA _________________________________________ Profº. Dr. Julio Cesar Pieczarka (Membro) _________________________________________ Profº. Dr. Pablo Suárez (Membro) _________________________________________ Profº. Dr. Leonardo dos Santos Sena (Membro) _________________________________________ Profº. Dr. Carlos Eduardo Matos Carvalho Bastos (suplente) Apresentado em 29 de Fevereiro de 2016 Belém – PA 2016 i “.. -

A New Species of Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from Rio Madeira Basin, Amazonas State, Brazil
Neotropical Ichthyology, 17(2): e180135, 2019 Journal homepage: www.scielo.br/ni DOI: 10.1590/1982-0224-20180135 Published online: 27 June 2019 (ISSN 1982-0224) Copyright © 2019 Sociedade Brasileira de Ictiologia Printed: 30 June 2019 (ISSN 1679-6225) Original article A new species of Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from rio Madeira basin, Amazonas State, Brazil Alessandro Gasparetto Bifi, Renildo Ribeiro de Oliveira and Lúcia Rapp Py-Daniel During the Mosaico do Apuí expedition in 2006, a new species of Ancistrus was found in the rio Sucunduri, a large tributary of the rio Madeira basin in south Amazonas State. The new species has a unique color pattern for the genus and is distinguished by the presence of alternate light and dark vertical bars on the trunk. Other Ancistrini genera have species with a similar color pattern, but the new species is clearly distinguished by a lack of plates on the snout and presence of fleshy tentacles on the snout as in allAncistrus representatives. We also provide comments on the taxonomic status of the A. bolivianus, A. heterorhynchus and A. marcapatae. Keywords: Ancistrini, Ancistrus bolivianus, Ancistrus heterorhynchus, Ancistrus marcapatae, Armored catfish. Durante a expedição ao mosaico do Apuí em 2006, uma nova espécie de Ancistrus foi coletada no rio Sucunduri, um grande tributário da bacia do rio Madeira, na região sul do estado do Amazonas. A nova espécie possui um padrão de colorido único para o gênero e é diagnosticada pela presença de faixas verticais claras no tronco intercaladas com faixas escuras. Outros gêneros em Ancistrini possuem espécies com um padrão de colorido similar, porém a espécie nova é claramente distinta pela ausência de placas e presença de tentáculos no focinho, conforme todos os representantes de Ancistrus. -

Astyanax Cavefish Bibliography, Chronological
Astyanax Cavefish Bibliography, chronological 552 citations from the Cave Life Bibliography William R. Elliott, [email protected] Hubbs, Carl L., and William T. Innes. 1936. The first known blind fish of the family Characidae: A new genus from Mexico. Occasional Papers of the Museum of Zoology, University of Michigan, no. 342. 7 pp., 1 pl. Muir, JM. 1936. Geology of the Tampico Region, Mexico. Special Volume ed. Tulsa, Oklahoma. American Association of Petroleum Geologists, Tulsa, 280 pp. Hykes, O.V. 1937. _Anoptichthys jordani_, Hubbs und Innes. Akvaristické listy, 11:108-109. Innes, William T. 1937. A cavern characin _Anoptichthys jordani_, Hubbs & Innes. Aquarium, Philadelphia, 5(10):200-202. Jordan, C. Basil. 1937. Bringing in the new cave fish _Anoptichthys jordani_ Hubbs and Innes. Aquarium, Philadelphia, 5(10):203-204. Anonymous. 1940. Expedición para recoger peces ciegos en México. Ciencia, 1:221. Bridges, William. 1940. The blind fish of La Cueva Chica. Bulletin of the New York Zoological Society. 43:74-97. De Buen, Fernando. 1940. Lista de peces de agua dulce de México. En preparación de su catálogo. Trabajos de Estación Limnológica de Pátzcuaro, 2. 66 pp. Gresser, E. B., and C. M. Breder, Jr. 1940. The histology of the eye of the cave characin, _Anoptichthys_. Zoologica, New York, 25(10):113-116, pls. I- III. Heim, A. 1940. The front ranges of the Sierra Madre Oriental, Mexico, from Ciudad Victoria to Tamazunchale. Eclogae Geolicae Helvetiae, 33:313-352. Breder, Charles M., Jr., and Edward B. Gresser. 1941. Correlations between structural eye defects and behavior in the Mexican blind characin.