Etude Comparã©E Du Comportement
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Preserving the Tree of Life of the Fish Family Cyprinidae in Africa in the Face of the Ongoing Extinction Crisis
Genome Preserving the tree of life of the fish family Cyprinidae in Africa in the face of the ongoing extinction crisis Journal: Genome Manuscript ID gen-2018-0023.R3 Manuscript Type: Article Date Submitted by the 02-Mar-2019 Author: Complete List of Authors: Adeoba, Mariam; University of Johannesburg Tesfamichael, Solomon; University of Johannesburg Yessoufou, Kowiyou; University of Johannesburg Conservation, African freshwater Ecosystems, IUCN Red List, EDGE, DNA Keyword: Draft barcoding Is the invited manuscript for consideration in a Special 7th International Barcode of Life Issue? : https://mc06.manuscriptcentral.com/genome-pubs Page 1 of 42 Genome Preserving the tree of life of the fish family Cyprinidae in Africa in the face of the ongoing extinction crisis Mariam Salami1, Solomon Tesfamichael2, Kowiyou Yessoufou2 1Department of Zoology, University of Johannesburg, Kingsway Campus, PO Box 524, Auckland Park 2006, South Africa 2Department of Geography, Environmental Management and Energy studies, University of Johannesburg, Kingsway Campus, PO Box 524, Auckland Park 2006, South Africa *Corresponding author: Kowiyou Yessoufou [email protected] Draft 1 https://mc06.manuscriptcentral.com/genome-pubs Genome Page 2 of 42 Abstract Our understanding of how the phylogenetic tree of fishes might be affected by the ongoing extinction risk is poor. This is due to the unavailability of comprehensive DNA data, especially for many African lineages. In addition, the ongoing taxonomic confusion within some lineages, e.g. Cyprinidae, makes it difficult to contribute to the debate on how the fish tree of life might be shaped by extinction. Here, we combine COI sequences and taxonomic information to assemble a fully sampled phylogeny of the African Cyprinidae and investigate whether we might lose more phylogenetic diversity (PD) than expected if currently-threatened species go extinct. -

Reproductive Behavior, Development and Eye Regression in the Cave
Neotropical Ichthyology, 7(3):479-490, 2009 Copyright © 2009 Sociedade Brasileira de Ictiologia Reproductive behavior, development and eye regression in the cave armored catfish, Ancistrus cryptophthalmus Reis, 1987 (Siluriformes: Loricariidae), breed in laboratory Sandro Secutti and Eleonora Trajano The troglobitic armored catfish, Ancistrus cryptophthalmus (Loricariidae, Ancistrinae) is known from four caves in the São Domingos karst area, upper rio Tocantins basin, Central Brazil. These populations differ in general body shape and degree of reduction of eyes and of pigmentation. The small Passa Três population (around 1,000 individuals) presents the most reduced eyes, which are not externally visible in adults. A small group of Passa Três catfish, one male and three females, reproduced spontaneously thrice in laboratory, at the end of summertime in 2000, 2003 and 2004. Herein we describe the reproductive behavior during the 2003 event, as well as the early development of the 2003 and 2004 offsprings, with focus on body growth and ontogenetic regression of eyes. The parental care by the male, which includes defense of the rock shelter where the egg clutch is laid, cleaning and oxygenation of eggs, is typical of many loricariids. On the other hand, the slow development, including delayed eye degeneration, low body growth rates and high estimated longevity (15 years or more) are characteristic of precocial, or K-selected, life cycles. In the absence of comparable data for close epigean relatives (Ancistrus spp.), it is not possible to establish whether these features are an autapomorphic specialization of the troglobitic A. cryptophthalmus or a plesiomorphic trait already present in the epigean ancestor, possibly favoring the adoption of the life in the food-poor cave environment. -

Checklist of Fish and Invertebrates Listed in the CITES Appendices
JOINTS NATURE \=^ CONSERVATION COMMITTEE Checklist of fish and mvertebrates Usted in the CITES appendices JNCC REPORT (SSN0963-«OStl JOINT NATURE CONSERVATION COMMITTEE Report distribution Report Number: No. 238 Contract Number/JNCC project number: F7 1-12-332 Date received: 9 June 1995 Report tide: Checklist of fish and invertebrates listed in the CITES appendices Contract tide: Revised Checklists of CITES species database Contractor: World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge, CB3 ODL Comments: A further fish and invertebrate edition in the Checklist series begun by NCC in 1979, revised and brought up to date with current CITES listings Restrictions: Distribution: JNCC report collection 2 copies Nature Conservancy Council for England, HQ, Library 1 copy Scottish Natural Heritage, HQ, Library 1 copy Countryside Council for Wales, HQ, Library 1 copy A T Smail, Copyright Libraries Agent, 100 Euston Road, London, NWl 2HQ 5 copies British Library, Legal Deposit Office, Boston Spa, Wetherby, West Yorkshire, LS23 7BQ 1 copy Chadwick-Healey Ltd, Cambridge Place, Cambridge, CB2 INR 1 copy BIOSIS UK, Garforth House, 54 Michlegate, York, YOl ILF 1 copy CITES Management and Scientific Authorities of EC Member States total 30 copies CITES Authorities, UK Dependencies total 13 copies CITES Secretariat 5 copies CITES Animals Committee chairman 1 copy European Commission DG Xl/D/2 1 copy World Conservation Monitoring Centre 20 copies TRAFFIC International 5 copies Animal Quarantine Station, Heathrow 1 copy Department of the Environment (GWD) 5 copies Foreign & Commonwealth Office (ESED) 1 copy HM Customs & Excise 3 copies M Bradley Taylor (ACPO) 1 copy ^\(\\ Joint Nature Conservation Committee Report No. -

A Guide to the Parasites of African Freshwater Fishes
A Guide to the Parasites of African Freshwater Fishes Edited by T. Scholz, M.P.M. Vanhove, N. Smit, Z. Jayasundera & M. Gelnar Volume 18 (2018) Chapter 2.1. FISH DIVERSITY AND ECOLOGY Martin REICHARD Diversity of fshes in Africa Fishes are the most taxonomically diverse group of vertebrates and Africa shares a large portion of this diversity. This is due to its rich geological history – being a part of Gondwana, it shares taxa with the Neotropical region, whereas recent close geographical affnity to Eurasia permitted faunal exchange with European and Asian taxa. At the same time, relative isolation and the complex climatic and geological history of Africa enabled major diversifcation within the continent. The taxonomic diversity of African freshwater fshes is associated with functional and ecological diversity. While freshwater habitats form a tiny fraction of the total surface of aquatic habitats compared with the marine environment, most teleost fsh diversity occurs in fresh waters. There are over 3,200 freshwater fsh species in Africa and it is likely several hundreds of species remain undescribed (Snoeks et al. 2011). This high diversity and endemism is likely mirrored in diversity and endemism of their parasites. African fsh diversity includes an ancient group of air-breathing lungfshes (Protopterus spp.). Other taxa are capable of breathing air and tolerate poor water quality, including several clariid catfshes (e.g., Clarias spp.; Fig. 2.1.1D) and anabantids (Ctenopoma spp.). Africa is also home to several bichir species (Polypterus spp.; Fig. 2.1.1A), an ancient fsh group endemic to Africa, and bonytongue Heterotis niloticus (Cuvier, 1829) (Osteoglossidae), a basal actinopterygian fsh. -

Volume 2. Animals
AC20 Doc. 8.5 Annex (English only/Seulement en anglais/Únicamente en inglés) REVIEW OF SIGNIFICANT TRADE ANALYSIS OF TRADE TRENDS WITH NOTES ON THE CONSERVATION STATUS OF SELECTED SPECIES Volume 2. Animals Prepared for the CITES Animals Committee, CITES Secretariat by the United Nations Environment Programme World Conservation Monitoring Centre JANUARY 2004 AC20 Doc. 8.5 – p. 3 Prepared and produced by: UNEP World Conservation Monitoring Centre, Cambridge, UK UNEP WORLD CONSERVATION MONITORING CENTRE (UNEP-WCMC) www.unep-wcmc.org The UNEP World Conservation Monitoring Centre is the biodiversity assessment and policy implementation arm of the United Nations Environment Programme, the world’s foremost intergovernmental environmental organisation. UNEP-WCMC aims to help decision-makers recognise the value of biodiversity to people everywhere, and to apply this knowledge to all that they do. The Centre’s challenge is to transform complex data into policy-relevant information, to build tools and systems for analysis and integration, and to support the needs of nations and the international community as they engage in joint programmes of action. UNEP-WCMC provides objective, scientifically rigorous products and services that include ecosystem assessments, support for implementation of environmental agreements, regional and global biodiversity information, research on threats and impacts, and development of future scenarios for the living world. Prepared for: The CITES Secretariat, Geneva A contribution to UNEP - The United Nations Environment Programme Printed by: UNEP World Conservation Monitoring Centre 219 Huntingdon Road, Cambridge CB3 0DL, UK © Copyright: UNEP World Conservation Monitoring Centre/CITES Secretariat The contents of this report do not necessarily reflect the views or policies of UNEP or contributory organisations. -
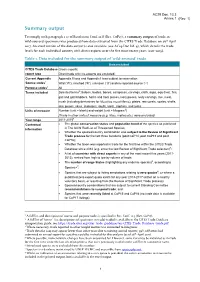
Summary Output
AC29 Doc. 13.3 Annex 1 Summary output To comply with paragraph 1 a) of Resolution Conf. 12.8 (Rev. CoP17), a summary output of trade in wild-sourced specimens was produced from data extracted from the CITES Trade Database on 26th April 2017. An excel version of the data output is also available (see AC29 Doc Inf. 4), which details the trade levels for each individual country with direct exports over the five most recent years (2011-2015). Table 1. Data included for the summary output of ‘wild-sourced’ trade Data included CITES Trade Database Gross exports; report type Direct trade only (re-exports are excluded) Current Appendix Appendix II taxa and Appendix I taxa subject to reservation Source codes1 Wild (‘W’), ranched (‘R’), unknown (‘U’) and no reported source (‘-’) Purpose codes1 All Terms included Selected terms2: baleen, bodies, bones, carapaces, carvings, cloth, eggs, egg (live), fins, gall and gall bladders, horns and horn pieces, ivory pieces, ivory carvings, live, meat, musk (including derivatives for Moschus moschiferus), plates, raw corals, scales, shells, skin pieces, skins, skeletons, skulls, teeth, trophies, and tusks. Units of measure Number (unit = blank) and weight (unit = kilogram3) [Trade in other units of measure (e.g. litres, metres etc.) were excluded] Year range 2011-20154 Contextual The global conservation status and population trend of the species as published information in The IUCN Red List of Threatened Species; Whether the species/country combination was subject to the Review of Significant Trade process for the last three iterations (post CoP14, post CoP15 and post CoP16); Whether the taxon was reported in trade for the first time within the CITES Trade Database since 2012 (e.g. -

Inventaire Rapide Des Zones Humides Représentatives En République Démocratique Du Congo
Inventaire Rapide des Zones Humides Représentatives en République Démocratique du Congo Editors and Major Contributors : Michele Thieme, Aurelie Shapiro, Alejandra Colom, Uli Schliewen, Nikolai Sindorf, Andre Kamdem Toham Supported by the Swiss Grant for Africa, Ramsar Convention Secretariat Table of Contents Executive Summary I. Introduction II. Approach and Methodology for Rapid Inventory of Wetland Priority Areas A. Aquatic Ecosystem Classification and Gap Analysis B. Human Use / Suitability Analysis C. Marxan Model D. Delineation and Prioritization of Wetland Areas E. Threats assessment, socio economic importance and key stakeholders Appendix I. Contributors Appendix II. List of Protected Areas used in the Gap Analysis Appendix III. Photos of Wetland Areas Appendix IV. List of Fish Species by Wetland Priority Area Executive Summary In 2002, the government of the Democratic Republic of Congo committed to increase its protected area coverage to 15% from just over 8%. WWF, RAMSAR, OSFAC and other partners supported the effort by convening an expert workshop in Kinshasa, DRC in November 2007, to undertake a country wide biodiversity assessment, including a rapid inventory and prioritization of the biodiversity value of the wetlands of the Democratic Republic of Congo. Several products were developed prior to the workshop to facilitate the work of the experts. These were: (1) a map classifying small sub‐basins in DRC by the predominant aquatic habitat types and (2) a map of human use/suitability. The aquatic habitat or ecosystem map was used as a basis for ensuring representation of all habitat types across the country and the suitability map provided a measure of the level of human impact. -

AC31 Doc. 13.4 A2
AC31 Doc.13.4 Annex 2 AC31 Doc. xx Annex X UNEP-WCMC Technical Report SELECTION OF SPECIES FOR INCLUSION IN THE REVIEW OF SIGNIFICANT TRADE FOLLOWING COP18: Extended Analysis AC31 Doc. 13.4 Annex 2 Selection of species for inclusion in the Review of Significant Trade following CoP18: Extended analysis Prepared for CITES Secretariat Published May 2020 Citation UNEP-WCMC. 2020. Selection of species for inclusion in the Review of Significant Trade following CoP18: Extended analysis. UNEP-WCMC, Cambridge. Copyright CITES Secretariat, 2020 The UN Environment Programme World Conservation Monitoring Centre (UNEP-WCMC) is a global Centre of excellence on biodiversity. The Centre operates as a collaboration between the UN Environment Programme and the UK-registered charity WCMC. Together we are confronting the global crisis facing nature. This publication may be reproduced for educational or non-profit purposes without special permission, provided acknowledgement to the source is made. Reuse of any figures is subject to permission from the original rights holders. No use of this publication may be made for resale or any other commercial purpose without permission in writing from the UN Environment Programme. Applications for permission, with a statement of purpose and extent of reproduction, should be sent to the Director, UNEP-WCMC, 219 Huntingdon Road, Cambridge, CB3 0DL, UK. The contents of this report do not necessarily reflect the views or policies of the UN Environment Programme, contributory organisations or editors. The designations employed and the presentations of material in this report do not imply the expression of any opinion whatsoever on the part of the UN Environment Programme or contributory organisations, editors or publishers concerning the legal status of any country, territory, city area or its authorities, or concerning the delimitation of its frontiers or boundaries or the designation of its name, frontiers or boundaries. -

Astyanax Cavefish Bibliography, Chronological
Astyanax Cavefish Bibliography, chronological 552 citations from the Cave Life Bibliography William R. Elliott, [email protected] Hubbs, Carl L., and William T. Innes. 1936. The first known blind fish of the family Characidae: A new genus from Mexico. Occasional Papers of the Museum of Zoology, University of Michigan, no. 342. 7 pp., 1 pl. Muir, JM. 1936. Geology of the Tampico Region, Mexico. Special Volume ed. Tulsa, Oklahoma. American Association of Petroleum Geologists, Tulsa, 280 pp. Hykes, O.V. 1937. _Anoptichthys jordani_, Hubbs und Innes. Akvaristické listy, 11:108-109. Innes, William T. 1937. A cavern characin _Anoptichthys jordani_, Hubbs & Innes. Aquarium, Philadelphia, 5(10):200-202. Jordan, C. Basil. 1937. Bringing in the new cave fish _Anoptichthys jordani_ Hubbs and Innes. Aquarium, Philadelphia, 5(10):203-204. Anonymous. 1940. Expedición para recoger peces ciegos en México. Ciencia, 1:221. Bridges, William. 1940. The blind fish of La Cueva Chica. Bulletin of the New York Zoological Society. 43:74-97. De Buen, Fernando. 1940. Lista de peces de agua dulce de México. En preparación de su catálogo. Trabajos de Estación Limnológica de Pátzcuaro, 2. 66 pp. Gresser, E. B., and C. M. Breder, Jr. 1940. The histology of the eye of the cave characin, _Anoptichthys_. Zoologica, New York, 25(10):113-116, pls. I- III. Heim, A. 1940. The front ranges of the Sierra Madre Oriental, Mexico, from Ciudad Victoria to Tamazunchale. Eclogae Geolicae Helvetiae, 33:313-352. Breder, Charles M., Jr., and Edward B. Gresser. 1941. Correlations between structural eye defects and behavior in the Mexican blind characin. -

Sixth International Conference of the Pan African Fish and Fisheries
SIXTH INTERNATIONAL CONFERENCE OF THE PAN AFRICAN FISH AND FISHERIES ASSOCIATION (PAFFA6) BOOK OF ABSTRACTS Sun N Sand Holiday Resort in Mangochi, Malawi 24th to 28th September 2018. “African Fish and Fisheries: Diversity, Conservation and Sustainable Management” About This Booklet This publication includes abstracts for oral presentations and poster presentations at the Sixth International Conference of The Pan African Fish And Fisheries Association (PAFFA6) held at Sun ‘n’ Sand Holiday Resort in Mangochi, Malawi from 24-28 September, 2018. Section One: Oral Presentations Oral presentations are grouped by conference theme. Please refer to the Conference Programme for details about date, time slot and location for each thematic session. Section Two: Poster Presentations Poster presentations are grouped by conference theme. Please refer to the Conference Programme for details about date, time slot, and location for group poster sessions. All presentations are subject to change after the printing of this publication. The 2018 PAFFA book of abstracts is sponsored by the Fisheries Integration of Society and Habitats Project (FISH) which is made possible by the generous support of the American people through the United States Agency for International Development (USAID) and implemented by Pact. "The contents, are the sole responsibility of LUANAR, Conference Organisers and Delegates and do not necessarily reflect the views of the FISH Project team and partners, USAID, or the United States Government (USG). 1 | P a g e “African Fish and Fisheries: Diversity, Conservation and Sustainable Management” KEY NOTE PRESENTATIONS – PLENARY SESSIONS (NYANJA HALL) Day 1, Monday, 24th September, 2018 Rapid Radiation of the Cichlids of Lake Malaŵi Jay R. -

The Status and Distribution of Freshwater Biodiversity in Central Africa
THE S THE STATUS AND DISTRIBUTION T A OF FRESHWATER BIODIVERSITY T U S IN CENTRAL AFRICA AND Brooks, E.G.E., Allen, D.J. and Darwall, W.R.T. D I st RIBU T ION OF F RE S HWA T ER B IODIVER S I T Y IN CEN CENTRAL AFRICA CENTRAL T RAL AFRICA INTERNATIONAL UNION FOR CONSERVATION OF NATURE WORLD HEADQUARTERS Rue Mauverney 28 1196 Gland Switzerland Tel: + 41 22 999 0000 Fax: + 41 22 999 0020 www.iucn.org/species www.iucnredlist.org The IUCN Red List of Threatened SpeciesTM Regional Assessment About IUCN IUCN Red List of Threatened Species™ – Regional Assessment IUCN, International Union for Conservation of Nature, helps the world find pragmatic solutions to our most pressing environment and development Africa challenges. The Status and Distribution of Freshwater Biodiversity in Eastern Africa. Compiled by William R.T. Darwall, Kevin IUCN works on biodiversity, climate change, energy, human livelihoods and greening the world economy by supporting scientific research, managing G. Smith, Thomas Lowe and Jean-Christophe Vié, 2005. field projects all over the world, and bringing governments, NGOs, the UN and companies together to develop policy, laws and best practice. The Status and Distribution of Freshwater Biodiversity in Southern Africa. Compiled by William R.T. Darwall, IUCN is the world’s oldest and largest global environmental organization, Kevin G. Smith, Denis Tweddle and Paul Skelton, 2009. with more than 1,000 government and NGO members and almost 11,000 volunteer experts in some 160 countries. IUCN’s work is supported by over The Status and Distribution of Freshwater Biodiversity in Western Africa. -

Download (2MB)
A translation of certain extracts from: "Les genres des poissons d'eau douce de l'Afrique" (The genera of freshwater fish from Africa) by M. Poll. (1957) Ann. Mus. Roy. Congo Belge. Ser. 8, Zool. 54: 1-191. Item Type monograph Authors Mann, M.J.; Poll, M. Publisher East African Freshwater Fisheries Research Organization Download date 25/09/2021 20:45:38 Link to Item http://hdl.handle.net/1834/32732 ~ ,_'?""W •••••.•• ~...-:-". =-...-=---.......-.~-'~.,~."._~~:'~~~:.,~~~'-'~ I. EAfFRO OCCAS~ONAL PAf',"f; L=LI.~ A translation of certain extracts from : "Les genres des poissons d I eau douce de:.a.' Afrique" (The genera of freshwater fish from Africa) by M. POLL (1957) o Ann. Mus. Roy. Congo Beige. Ser.S , Zool • -..2.1' 1-191 translated by M.J. Mann (Fisheries Research Officer) E.A. F.F.R. O. P.O. Box 343, Jinja,, Uganda. December, 1967. CONTENTS (Seotions "GYpedin lower oase are not inoluded in the translation) TR.AN"S.LA.TOR'S INTB.ODUCTION ••••••••••• '. '••• '•• '•••••• '•• '•• '•• '•••• '••• '. -•••• '" ••.••• I Il'fffiODUCTION ••••••••••• '.,•• '••.•.•.• 0 .' •• '.' ••• ' ••••••••••• '.' •••••••••••• ,., •• ' ••••• ".'.'., •• " 1 General table of the classes, order~, t'amiliesandgenera of fish represented in the freshwater of Afrioa General oharaoters of each family "' THE GENERAL GEOGRAPHICAL DISTRIBUTION OF THE AFRICAN FISH FAUNA •••,••••;;. 3 THE GEOGRAPHICAL DISTRIBUTION OF FRESHWATER FISHES IN THE CONGO BASIN; •••12 THE DIAGNOSTIC CHARACTERS OF FISH ••••••••••.• ~•••••• ,•• ~.•'••• -" ••••••••••••• 17 KEY FOR THE IDENTIFICATION OF THE FAMILIES OF AFRICAN FRESHWATER FISH ••••18 KEY FOR THE IDENTIFICATION OF THE GENERA OF AFRICAN FRESHWATER FISH ••••••21 Bibliography Index of figures Index of soientifio names Map of Afrioan iohthyologioal regions • I Translator's introduction At present there is no convenient key in English for the identification of the species of freshwater fish from the East African political region which includes Kenya, Tanzania and Uganda.